
Войти
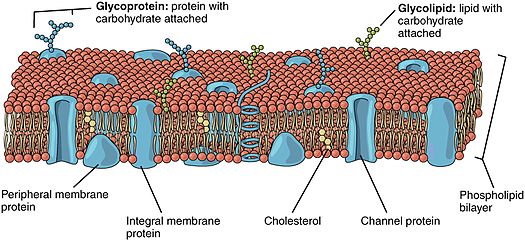 Липидная мембрана с различными белками
Липидная мембрана с различными белками Липид-заякоренные белки (также известные как липид-связанные белки ) - белки, расположенные на поверхности клеточной мембраны, которые ковалентно присоединены к липидам, встроенным в клеточную мембрану. Эти белки вставляются и занимают место в двухслойной структуре мембраны вместе с аналогичными хвостами жирных кислот. Заякоренный липидом белок может располагаться по обе стороны от клеточной мембраны. Таким образом, липид служит для закрепления белка на клеточной мембране. Они представляют собой тип протеолипидов.
. Липидные группы играют роль во взаимодействии с белками и могут вносить вклад в функцию белка, к которому они присоединены. Кроме того, липид служит медиатором мембранных ассоциаций или детерминантом специфических белок-белковых взаимодействий. Например, липидные группы могут играть важную роль в повышении молекулярной гидрофобности. Это обеспечивает взаимодействие белков с клеточными мембранами и доменами белков. В динамической роли липидирование может изолировать белок от его субстрата, чтобы инактивировать белок, а затем активировать его посредством презентации субстрата.
В целом существует три основных типа липидно-заякоренных белков, которые включают пренилированные белки., жирные ацилированные белки и гликозилфосфатидилинозитол-связанные белки (GPI) . Белок может иметь несколько липидных групп, ковалентно связанных с ним, но место, где липид связывается с белком, зависит как от липидной группы, так и от белка.
 Изопреновая единица
Изопреновая единица Пренилированные белки представляют собой белки с ковалентно присоединенными гидрофобными изопреновыми полимерами (т.е. разветвленными пятиуглеродными углеводородами) у цистеиновых остатков белка. Более конкретно, эти изопреноидные группы, обычно фарнезил (15-углеродный) и геранилгеранил (20-углеродный), присоединены к белку через тиоэфирные связи на остатках цистеина вблизи С-конца белок. Это пренилирование липидных цепей до белков облегчает их взаимодействие с клеточной мембраной.
 Caax Box
Caax Box Мотив пренилирования «CAAX box» является наиболее распространенным сайтом пренилирования в белках, то есть сайтом где ковалентно присоединены фарнезил или геранилгеранил. В последовательности CAAX-бокса C представляет собой пренилированный цистеин, A представляет любую алифатическую аминокислоту, а X определяет тип пренилирования, которое будет происходить. Если X представляет собой Ala, Met, Ser или Gln, белок будет фарнезилироваться с помощью фермента фарнезилтрансфераза, а если X представляет собой Leu, то белок будет геранилгеранилирован с помощью геранилгеранилтрансферазы I фермент. Оба эти фермента похожи, каждый из них содержит две субъединицы.
 Цепи пренилирования (например, геранилпирофосфат )
Цепи пренилирования (например, геранилпирофосфат )Пренилированные белки особенно важны для роста, дифференцировки и морфологии эукариотических клеток. Кроме того, пренилирование белков представляет собой обратимую посттрансляционную модификацию клеточной мембраны. Это динамическое взаимодействие пренилированных белков с клеточной мембраной важно для их сигнальных функций и часто не регулируется при таких болезненных процессах, как рак. Более конкретно, Ras - это белок, который подвергается пренилированию с помощью фарнезилтрансферазы, и при его включении он может включать гены, участвующие в росте и дифференцировке клеток. Таким образом, чрезмерная активация передачи сигналов Ras может привести к раку. Понимание этих пренилированных белков и их механизмы были важны для усилий по разработке лекарств для борьбы с раком.Другие пренилированные белки включают члены Rab и Rh o семейства, а также ламины.
Некоторые важные цепи пренилирования, которые участвуют в HMG-CoA редуктазе метаболическом пути, - это геранилгераниол, фарнезол и долихол. Эти изопреновые полимеры (например, геранилпирофосфат и фарнезилпирофосфат ) участвуют в конденсации через ферменты, такие как пренилтрансфераза, которые в конечном итоге циклизуются с образованием холестерина.
Жирные ацилированные белки представляют собой белки, которые были посттрансляционно модифицированы с целью включения ковалентного присоединения жирных кислот к определенным аминокислотным остаткам. Наиболее распространенными жирными кислотами, которые ковалентно присоединены к белку, являются насыщенная миристиновая (14-углеродная) кислота и пальмитиновая кислота (16-углеродная). Белки можно модифицировать, чтобы они содержали одну или обе эти жирные кислоты.
 Миристоилирование
Миристоилирование N-миристоилирование (то есть присоединение миристиновой кислоты), как правило, является необратимой модификацией белка, которая обычно происходит во время синтеза белка, в котором миризитовая кислота присоединена к α-аминогруппе N-концевого остатка глицина через амидную связь. Этой реакции способствует N-миристоилтрансфераза. Эти белки обычно начинаются с последовательности Met - Gly и с серина или треонина в положении 5. Белки, которые были миристоилированы, участвуют в каскад передачи сигналов, белок-белковые взаимодействия и механизмы, регулирующие нацеливание и функцию белков. Примером, в котором важно миристоилирование белка, является апоптоз, запрограммированная гибель клеток. После того, как белок BH3-агонист гибели взаимодействующего домена (Bid) был миристоилирован, он нацеливает белок на перемещение к митохондриальной мембране для высвобождения цитохрома с, что в конечном итоге приводит к гибели клетки. Другие миристоилированные белки, участвующие в регуляции апоптоза, - это актин и гельсолин.
 пальмитоилирование
пальмитоилирование S-пальмитоилирование (то есть присоединение пальмитиновой кислоты) обратимая модификация белка, при которой пальмитиновая кислота присоединяется к определенному остатку цистеина через связь тиоэфира. Термин S-ацилирование может также использоваться, когда другие средние и длинные цепи жирных кислот также присоединены к пальмитоилированным белкам. Консенсусной последовательности пальмитоилирования белка не обнаружено. Пальмитоилированные белки в основном обнаруживаются на цитоплазматической стороне плазматической мембраны, где они играют роль в трансмембранной передаче сигналов. Пальмитоильная группа может быть удалена пальмитоилтиоэстеразой. Считается, что это обратное пальмитоилирование может регулировать взаимодействие белка с мембраной и, таким образом, играть роль в процессах передачи сигналов. Кроме того, это позволяет регулировать субклеточную локализацию, стабильность и доставку белка. Примером, в котором пальмитоилирование белка играет роль в сигнальных путях клетки, является кластеризация белков в синапсе. Когда белок постсинаптической плотности 95 (PSD-95) пальмитоилирован, он ограничивается мембраной и позволяет ему связываться и кластеризовать ионные каналы в постсинаптической мембране. Таким образом, пальмитоилирование может играть роль в регуляции высвобождения нейромедиаторов.
Пальмитоилирование опосредует сродство белка к липидным рафтам и облегчает кластеризацию белков. Кластеризация может увеличить близость двух молекул. Альтернативно, кластеризация может изолировать белок от субстрата. Например, пальмитоилирование фосфолипазы D (PLD) изолирует фермент от его субстрата фосфатидилхолина. Когда уровни холестерина снижаются или уровни PIP2 увеличиваются, опосредованная пальмитатом локализация нарушается, фермент переходит в PIP2, где он встречает свой субстрат и активен посредством презентации субстрата.
 Структура якорь гликофосфатидилинозитола в плазматической мембране эукариотической клетки
Структура якорь гликофосфатидилинозитола в плазматической мембране эукариотической клетки Гликозилфосфатидилинозитол (GPI) белки присоединены к комплексной молекулярной группе GPI через амидную связь с белками C-концевая карбоксильная группа. Этот комплекс GPI состоит из нескольких основных компонентов, которые связаны между собой: фосфоэтаноламин, линейный тетрасахарид (состоящий из трех маннозы и глюкозаминила) и фосфатидилинозитол. Группа фосфатидилинозитола гликозидно связана с не-N-ацетилированным глюкозамином тетрасахарида. Затем образуется фосфодиэфирная связь между маннозой на невосстанавливающем конце (тетрасахарида) и фосфоэтаноламином. Затем фосфоэтаноламин связывается амидом с С-концом карбоксильной группы соответствующего белка. Присоединение GPI происходит за счет действия комплекса GPI-трансамидаза. Цепи жирных кислот фосфатидилинозита вставляются в мембрану и, таким образом, являются тем, что закрепляет белок на мембране. Эти белки расположены только на внешней поверхности плазматической мембраны.
Остатки сахара в тетрасахариде и остатки жирных кислот в группе фосфатидилинозит варьируются в зависимости от белка. Это большое разнообразие позволяет белкам GPI выполнять широкий спектр функций, в том числе действовать как гидролитические ферменты, молекула адгезии, рецепторы, ингибитор протеазы и регулятор комплемента. белки. Кроме того, белки GPI играют важную роль в эмбриогенезе, развитии, нейрогенезе, иммунной системе и оплодотворении. Более конкретно, белок GPI IZUMO1R / JUNO (названный в честь римской богини плодородия ) в плазме яиц играет важную роль в слиянии сперматозоидов. Высвобождение белка IZUMO1R / JUNO GPI из плазматической мембраны яйца не позволяет сперматозоиду слиться с яйцеклеткой, и предполагается, что этот механизм может способствовать блоку полиспермии на плазматической мембране в яйцах. Другие роли, которые допускает модификация GPI, связаны с мембранными микродоменами, временной гомодимеризацией или апикальной сортировкой в поляризованных клетках.