
Войти
| Кальциевый канал, зависящий от напряжения | |
|---|---|
 Кристаллографическая структура Кристаллографическая структура | |
| Идентификаторы | |
| Символ | Кальциевый канал, потенциалозависимый |
 Иммуногистохимический анализ кальциевого канала L-типа Cav1.3 (CACNA1D) в коре надпочечников человека. Заметная иммунореактивность была обнаружена в клубочковой зоне. На рисунке: ZG = клубочковая зона, ZF = фасцикулярная зона, AC = капсула надпочечника. Иммуногистохимия проводилась в соответствии с опубликованными методами.
Иммуногистохимический анализ кальциевого канала L-типа Cav1.3 (CACNA1D) в коре надпочечников человека. Заметная иммунореактивность была обнаружена в клубочковой зоне. На рисунке: ZG = клубочковая зона, ZF = фасцикулярная зона, AC = капсула надпочечника. Иммуногистохимия проводилась в соответствии с опубликованными методами.  Кальциевый канал L-типа, субъединицы которого помечены вместе с некоторыми лекарствами, которые, как известно, ингибируют канал.
Кальциевый канал L-типа, субъединицы которого помечены вместе с некоторыми лекарствами, которые, как известно, ингибируют канал. Кальциевый канал L-типа (также известный как дигидропиридиновый канал или канал DHP ) является частью высоковольтного активированного семейства зависимых от напряжения кальциевых каналов. «L» означает длительный, что означает продолжительность активации. Этот канал состоит из четырех субъединиц (Cav1.1, Cav1.2, Cav1.3, Cav1.4 ).
кальциевые каналы L-типа отвечают за возбуждение-сокращение сцепление скелетной, гладкой, сердечной мышцы и для секреции альдостерона в эндокринных клетках коры надпочечников. Они также обнаружены в нейронах и с помощью кальциевых каналов L-типа в эндокринных клетках регулируют нейрогормоны и нейротрансмиттеры. Также было замечено, что они играют роль в экспрессии генов, стабильности мРНК, выживаемости нейронов, ишемическом повреждении аксонов, синаптической эффективности, а также в активации и деактивации других ионных каналов.
В сердечных миоцитах L Кальциевый канал -типа пропускает внутрь ток Са и запускает высвобождение кальция из саркоплазматического ретикулума, активируя рианодиновый рецептор 2 (RyR2) (кальций-индуцированное высвобождение кальция). Фосфорилирование этих каналов увеличивает их проницаемость для кальция и увеличивает сократительную способность соответствующих сердечных миоцитов.
L-тип блокаторы кальциевых каналов препараты используются в качестве сердечных антиаритмических или гипотензивных, в зависимости от того, имеют ли препараты более высокое сродство к сердце (фенилалкиламины, например верапамил ) или для кровеносных сосудов (дигидропиридины, например нифедипин ).
В в скелетных мышцах очень высокая концентрация кальциевых каналов L-типа, расположенных в Т-канальцах. Деполяризация мышц приводит к большим токам затвора, но аномально низкому потоку кальция, что теперь объясняется очень медленная активация ионных токов. По этой причине мало или совсем не проходит Са через мембрану Т-канальца во время одного потенциала действия.
В 1953 году Пол Фатт и Бернард Кац обнаружил потенциалзависимые кальциевые каналы в голени тейбовая мышца. Каналы демонстрировали различное напряжение активации и свойства проводимости кальция и, таким образом, были разделены на каналы активации высокого напряжения (HVA) и каналы активации низкого напряжения (LVA). После дополнительных экспериментов было обнаружено, что каналы HVA открываются для 1,4-дигидропиридина (DHP ). Используя DHP, они обнаружили, что каналы HVA специфичны для определенных тканей и реагируют по-разному, что привело к дальнейшей классификации каналов HVA на L-тип, P-тип и N-тип <3.>. Кальциевые каналы L-типа были секвенированы пептидами, и было обнаружено, что существует 4 типа кальциевых каналов L-типа: α 1 S (скелетные мышцы), α 1 C (сердечные), α 1 D (обнаружено в головном мозге) и α 1 F (обнаружено в сетчатке). В 2000 году, после того, как были проведены дополнительные исследования субъединиц α 1 в потенциал-управляемых кальциевых каналах, была использована новая номенклатура, которая назвала кальциевые каналы L-типа CaV1, а его субъединицы были названы CaV1.1, CaV1.2., CaV1,3 и CaV1,4. Исследования субъединиц CaV1 продолжают раскрывать больше информации об их структуре, функциях и фармацевтических применениях.
Кальциевые каналы L-типа содержат 5 различных субъединиц, α1 (170–240 кДа), субъединицы α2 (150 кДа), δ (17-25 кДа), β (50-78 кДа) и γ (32 кДа). Субъединицы α2, δ и β нековалентно связаны с субъединицей α1 и модулируют перенос ионов и биофизические свойства субъединицы α1. Субъединицы α2 и δ находятся во внеклеточном пространстве, а субъединицы β и γ расположены в цитозольном пространстве.
Субъединица α1 представляет собой гетеротетрамер, который имеет четыре трансмембранных области, известных как домены I-IV, которые шесть раз пересекают плазму как α-спирали, называемые S0-S6 (S0 и S1 вместе пересекают мембрану один раз). Субъединица α1 в целом содержит область измерения напряжения, пору проводимости и стробирующий аппарат. Как и большинство потенциал-управляемых ионных каналов, α-субъединица состоит из 4 субъединиц. Каждая субъединица образована 6 альфа-спиральными трансмембранными доменами, пересекающими мембрану (пронумерованные S1-S6). Субблоки S1-S4 составляют датчик напряжения, а субблоки S5-S6 составляют фильтр селективности. Чтобы воспринимать напряжение клетки, спирали S1-S3 содержат много отрицательно заряженных аминокислот, а спирали S4 содержат в основном положительно заряженные аминокислоты с P-петлей, соединяющей спирали S4 и S5. После доменов S1-6 есть шесть доменов C, которые состоят из двух EF-мотивов (C1-2 и C3-4) и домена Pre-IQ (C5) и домена IQ. (С6). На N-конце также есть два мотива EF-руки. Оба N- и C-конца находятся в цитозольном пространстве, причем C-конец намного длиннее, чем N-конец.
Известно, что субъединица β имеет четыре изоформы (β1-β4) для регулирования функций канала и связан с α1 через линкер α1 I и II в цитозоле в β α1- связывающем кармане (ABP). Каждая изоформа содержит домен гомологии src 3 (SH3) и гуанилаткиназоподобный домиан (GK), которые разделены доменом HOOK, а также три неструктурированных области.
α2 и δ субъединицы связаны между собой дисульфидными связями (иногда называемыми субъединицами α2δ) и взаимодействуют с α1. у них есть четыре известные изоформы, называемые от α2δ-1 до α2δ-2, и они содержат домен фон Виллебранда A (VWA) и домен кэша. Область α2 находится во внеклеточном пространстве, в то время как область δ находится в клеточной мембране, и было замечено, что она заякорена с помощью гликозилфосфатидилинозитола (GPI) якоря.
Субъединица γ имеет восемь изоформ (γ1-γ8) и связан с субъединицей α1 и обнаружен только в мышечных клетках в каналах CaV1.1 и CaV1.2. О субъединице γ известно немного, но она связана с взаимодействиями в гидрофобных силах.
Открытие поры в кальциевых каналах L-типа происходит в субъединице α1. Когда мембрана деполяризуется, спираль S4 перемещается через линкеры S4 и S5 к цитоплазматическим концам спиралей S5 и S6. Это открывает ворота активации, которые образованы внутренней стороной спиралей S6 в субъединице α1.
Наиболее распространенным способом аутоингибирования кальциевых каналов L-типа является Са + 2 / Кулачковый комплекс. Когда поры открываются и вызывает приток кальция, кальций связывается с кальмодулином, а затем взаимодействует с петлей, которая соединяет соседние мотивы EF-руки и вызывает конформационные изменения в EF- мотив руки, чтобы он взаимодействовал с порой, вызывая быстрое торможение в канале. До сих пор ведутся споры о том, где и как взаимодействуют поры и EF-рука. Гидрофобные карманы в комплексе Ca + 2 / Cam также будут связываться с тремя участками домена IQ, известными как «ароматические якоря». Комплекс CA + 2 / Cam имеет высокое сродство к кальциевым каналам L-типа, что позволяет ему блокироваться, даже когда в клетке присутствует небольшое количество кальция. Пора в конечном итоге закрывается по мере реполяризации клетки и вызывает конформационные изменения в канале, переводя ее в закрытую конформацию.
Одной из наиболее известных характеристик кальциевых каналов L-типа является его уникальная чувствительность к 1,4-дигидропиридинам (DHP). В отличие от других потенциалзависимых кальциевых каналов, кальциевые каналы L-типа устойчивы к препаратам, ингибирующим -CT X (GVIA) и ⍵-AG A (IVA).
Хорошо наблюдаемая форма модуляция обусловлена альтернативным соединением. Распространенной формой модуляции от альтернативного сплайсинга является C-концевой модулятор (CTM). Он имеет положительно заряженную α-спираль на С-конце, называемую DCRD, и отрицательно заряженную спираль сразу после мотива IQ (сайт взаимодействия CaM), называемую PCRD. Две спирали могут образовывать структуру, которая конкурентно связывается с CaM, чтобы снизить вероятность открытого состояния и более низкое кальций-зависимое ингибирование (CDI).
Альтернативное сплайсинг также наблюдается на субъединицах β для создания различных изоформ для придания каналам разных свойств из-за пальмитоилирования и. Другие формы модуляции β-субъединицы включают увеличение или уменьшение экспрессии субъединицы. Это связано с тем, что субъединицы β увеличивают вероятность открытия канала, активность в плазматической мембране и противодействуют убиквитинированию канала.
Кальциевые каналы L-типа являются также модулируется рецепторами, связанными с G-белком и адренергической нервной системой. Протеинкиназа A (PKA), активированная каскадом G-протеиновых рецепторов, может фосфорилировать L Кальциевые каналы -типа после того, как каналы образуют сигнальный комплекс с белками, заякоренными в А-киназе (AKAP), для увеличения кальциевого тока через канал, увеличения вероятности открытого состояния и ускоренного периода восстановления. Активированная фосфолипаза C (PLC) из рецепторов, связанных с G-белком, может разрушать полифосфоинозитиды, уменьшая ток кальция в каналах на 20-30%.
Было замечено, что адренергическая нервная система модулирует L кальциевые каналы -типа путем отщепления С-концевого фрагмента, когда β-адренорецептор стимулируется для увеличения активации каналов.
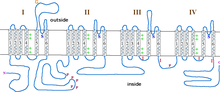 Альфа-субъединица общего потенциалзависимого ионного канала
Альфа-субъединица общего потенциалзависимого ионного канала Эта статья включает текст из Национальной медицинской библиотеки США, которая находится в общественном достоянии.
.