
Войти
Для рака, вторжение - это прямое распространение и проникновение раковые клетки в соседние ткани. Его обычно отличаются от метастазов, которые представляют собой распространение раковых клеток через систему кровообращения или лимфатическую систему в более отдаленные места. Тем не менее, лимфоваскулярная инвазия обычно является первым этапом метастазирования.
Многочисленные исследования подтвердили существование двух основных моделей инвазии раковых клеток посредством передачи клеток : коллективная миграция клеток и миграция отдельных клеток, в результате которых опухоль клетки переносят барьеры внеклеточного матрикса и распространяются в окружающие ткани. Механически-генетические механизмы, лежащие в основе генетически-генетических механизмов. Два типа мигающих опухолевых клеток, мезенхимальные (фибробластоподобные) и амебоидные, наблюдаются в каждом типе инвазии раковых клеток. В этом обзоре представлены различные варианты между методами использования раковых клеток, роль эпителиально-мезенхимальных, коллективно-амебоидных, мезенхимально-амебоидных и амебоид-мезенхимальных переходов, а также различных опухолевых факторов и стромальных молекул в опухолевой инвазии.. Собранные данные и факты важны для понимания того, как характер инвазии раковых клеток вызывает прогрессирование рака и эффективной терапии. Приведены убедительные доказательства того, что морфологические проявления паттернов инвазии характеризуются разнообразием тканевых (опухолевых) структур. Представлены результаты наших собственных исследований, чтобы показать связь прогрессирования рака груди с использованием видов клеточной адгезии в опухолевых клетках. различных морфологических структур.
Результаты испытаний экспериментальных и клинических испытаний злокачественных новообразований показали, что инвазивный рост и метастазирование проявления проявлениями прогрессирования опухоли, которые имеют два связанных процесса.
Злокачественная опухоль проявляет реализацию такого биологического явления, как метастатический каскад это уникальная многоступенчатая «программа», в которой инвазия клеток является спусковым крючком и способствует прогрессированию рака и метастазирования в отдаленные органы и ткани. Массивные метастатические болезни приводят к развитию тяжелой органной недостаточности и, как следствие, к смерти пациента. Диапазон между «конечными» точками сложного инвазивного метастатического процесса - вторжением первичной опухоли в окружающие ткани и образование метастатических очагов - состоит из нескольких этапов, прохождение которых строго необходимо для успешного развития и последующего прогрессирования опухолевого роста. : выживаемость и присутствие в системном кровотоке, экстравазация с помощью колонизацией органов опухолевыми клетками и образование клинически определяемых метастазов. Внеклеточное матрикса, как тканевое микроокружение, сохраненное за счет увеличения давления на опухолевые клетки. Факторы, ограничивающие рост злокачественного новообразования, включают базальную мембрану и различные компоненты окружающей среды, повышенное интерстициальное давление, ограниченное поступление кислорода к опухолевым клеткам и образование активных форм кислорода, условия гипоксии и постоянное воздействие на клетки иммунной системы. Противоречивые клетки, противодействующие факторам микроокружения, приобретают агрессивный фенотип и способность к метастатической прогрессии, подвергаются внутриопухолевой гетерогенности, могут подвергнуться регрессии и гибели, тогда как другие клетки, противостоящие, мощные факторы микроокружения. Инвазивный рост опухоли возможен за счет улучшения качества клеток от опухолевой массы за счет уменьшения или полной потери молекул межклеточной адгезии, и, следовательно, клетки приобретают способность к аномально высокой подвижности, позволяя проникать через жесткие структурные элементы окружающей строма. В этом процессе в процессе инвазии во многом участвуют различные молекулярные и клеточные механизмы, которые, согласно данным, зависят от других биологических явлений - эпителиально-мезенхимальной трансформации, впервые описанной E.D. Сено в 1995 году. Позднее термин «эпителиально-мезенхимальный переход» (EMT) был использован, чтобы прояснить обратимость этого процесса. В настоящее время известно, что ЭМП лежит в основе процессов эмбриогенеза, воспаления и регенерации тканей и безусловно играет ключевую роль в механизме канцерогенеза.
Опухолевые клетки Распространение в окружающих ткани и отдаленные органы, как известно, воспроизводит механизмы и типы миграции, характерные для нормальных, неопухолевых клеток во время физиологических процессов. Опухолевые клетки, аналогичные нормальным клеткам, способны активировать эти механизмы для своей собственной формы создания условий для движения, а также ремоделирования окружающих тканей для формирования путей перемещения. Основная проблема заключается в том, что опухолевые клетки, в отличие от нормальных, не физиологических «стоп-сигналов» для прекращения этих процессов. Скорее всего, это приводит к установлению миграции и прогрессированию опухоли.
Было установлено, что злокачественные клетки используют встроенные генетические программы для реализации процессов, которые определяют инвазивный рост и возможность метастазов. Например, движение индивидуальной клетки наблюдается во время эмбрионального развития и воспаления (например, миграция лейкоцитов). Подобный механизм распространения типичен для раковых клеток во время прогрессирования и метастазирования опухоли.
Наряду с миграцией отдельных клеток, коллективная миграция клеток может происходить, когда группы прочно связаны между собой опухолевых клеток, мигрирующих. Этот тип движения на перестройку тканей, основанный на процессах эмбрионального морфогенеза, является важным компонентом заживления раневых поверхностей.
Таким образом, ключевой моментом является то, что злокачественные опухолевые клетки используют широко используемые механизмы обоих коллективных механизмов. как физиологические прототипы в процессе инвазивного роста и метастазирования.
В настоящее время, основные данные комплекса определенных морфологических и молекулярно-генетических параметров, два принципиально разных Различают паттерны инвазивного роста: групповая (групповая) миграция клеток и миграция одиночных клеток (индивидуальная) миграция: рис. 1). При этом тип миграции во многом определяется особенностями тканевого микроокружения и зависит от молекулярных изменений в опухолевых клетках.
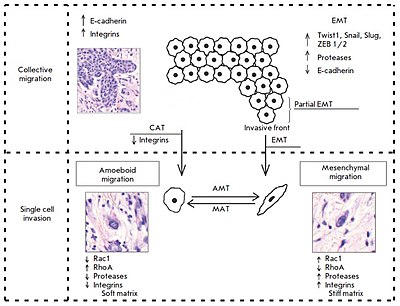 Паттерны инвазии раковых клеток: коллективная и индивидуальная миграция клеток. При коллективной экспрессии клеток опухолевые клетки E-экспрессию E-экспрессию. Эпителиально-мезенхимальные (EMT) и коллективно-амебоидные (CAT) переходы являются тригером между коллективной инвазией клеток и миграцией отдельных клеток. EMT включает активацию таких факторов транскрипции, как TWIST1, Snail, Slug, ZEB1 / 2, снижение экспрессии E-кадгерина и повышение активности протеазы. Во время ЭМП опухолевые клетки приобретают мезенхимальный фенотип, отделяются от опухолевой массы и мигрируют по мезенхимальному механизму. Напротив, частичная ЭМП, специфичная для инвазивного фронта опухоли, означает, что опухолевые клетки сохраняют межклеточную адгезию, но уже обладают миграционной способностью. Этот фенотиполевых клеток был назван «эпителиально-мезенхимальным» фенотипом. При CAT, которая имеет место, когда интегрины β1 подавляются, опухолевые клетки отделяются от опухолевой массы и перемещаются по амебоидному механизму. Амебоидная миграция включает снижение экспрессии протеазы и интегрина и изменения активности GTPases - ингибирование Rac1 и активацию RhoA. Этот тип движения происходит в рыхлом / мягком внеклеточном матриксе. Напротив, мезенхимальная миграция связана с противоположным фенотипом и преобладает в плотном / жестком матриксе. Эти два типа движения очень пластичны и могут преобразовываться друг в друга в зависимости от типа внеклеточного матрикса и внутриклеточной регуляции. Таким образом, предполагаются мезенхимально-амебоидный (MAT) и амебоид-мезенхимальный (AMT) переходы.
Паттерны инвазии раковых клеток: коллективная и индивидуальная миграция клеток. При коллективной экспрессии клеток опухолевые клетки E-экспрессию E-экспрессию. Эпителиально-мезенхимальные (EMT) и коллективно-амебоидные (CAT) переходы являются тригером между коллективной инвазией клеток и миграцией отдельных клеток. EMT включает активацию таких факторов транскрипции, как TWIST1, Snail, Slug, ZEB1 / 2, снижение экспрессии E-кадгерина и повышение активности протеазы. Во время ЭМП опухолевые клетки приобретают мезенхимальный фенотип, отделяются от опухолевой массы и мигрируют по мезенхимальному механизму. Напротив, частичная ЭМП, специфичная для инвазивного фронта опухоли, означает, что опухолевые клетки сохраняют межклеточную адгезию, но уже обладают миграционной способностью. Этот фенотиполевых клеток был назван «эпителиально-мезенхимальным» фенотипом. При CAT, которая имеет место, когда интегрины β1 подавляются, опухолевые клетки отделяются от опухолевой массы и перемещаются по амебоидному механизму. Амебоидная миграция включает снижение экспрессии протеазы и интегрина и изменения активности GTPases - ингибирование Rac1 и активацию RhoA. Этот тип движения происходит в рыхлом / мягком внеклеточном матриксе. Напротив, мезенхимальная миграция связана с противоположным фенотипом и преобладает в плотном / жестком матриксе. Эти два типа движения очень пластичны и могут преобразовываться друг в друга в зависимости от типа внеклеточного матрикса и внутриклеточной регуляции. Таким образом, предполагаются мезенхимально-амебоидный (MAT) и амебоид-мезенхимальный (AMT) переходы. Определение механизма инвазии, используемого одиночными мигрирующими механизмами во время передачи данных. К сожалению, исследования, посвященные этой проблеме на молекулярном и морфологическом уровнях, в основном они проводились in vitro с использованием определенных клеточных линий.
Однако в настоящее время наблюдается значительный рост количества исследований, демонстрирующих интерес к исследованиям молекулярно-генетических опухолевых клеток, которые определяют основные различия между мезенхимальным и амебоидным типами клеток во время индивидуальной миграции, а также коллективной передачи.
Коллективная миграция
Коллективная миграция миграцией целых групп клеток, связанных между собой молекулами адгезии и другими коммуникационными узлами (рис. 1). Это основные функции этого типа инвазий, поскольку лежащие в основе клеточные механизмы являются теми же ключевыми ключевыми факторами, которые определяют основные степени миграции отдельных клеток.
Коллективная миграция клеток наблюдается при развитии и прогрессировании рака и эндометрия., рак простаты, колоректальный рак, крупноклеточная карцинома легкого, рабдомиосаркома, мелана, также большинство плоскоклеточных карцином.
В случае коллективной передачи раковые клетки, являющиеся частью опухолевой массы или отделяющиеся от нее в виде многоклеточных групп проникают в окружающие ткани и образуют короткие короткие хорды, скопления, полосы и широкие поля, а также структуры с просветом, что указывает на широкий спектр структурных элементов, участвующих в инвазии опухоли.
Как уже упоминается, коллективная миграция проявляется миграцией целых групп клеток, связанных между собой кадгеринами и межклеточными щелевыми соединениями. Движущаяся группа клеток имеет «передний край» или «передний фронт», который использует интегрины и протеазы (Рис. 1). Исследователи указывают на четкие различия в экспрессии генов и морфологии между «лидерными» клетками, образующими передний край, и «ведомыми» клетками, которые расположены позади них, на «заднем крае». «Лидеры» в клетках часто представляют собой мезенхимные трубчатые структуры, характеризующиеся меньшей упорядоченностью и структурной организацией, в то время как «последователи» имеют тенденцию образовывать более плотно упакованные розеткообразные трубчатые структуры с плотными межклеточными контактами.
В случае коллективной миграции опухолевые клетки образуют выступы (псевдоподии) на переднем крае, используют интегрины для формирования очаговых контактов с актиновым цитоскелетом и осуществляют протеолитическую деграда внеклеточного матрикса, создавая пространство для инвазии опухолевой ткани. и активное вовлечение в процесс сократительного аппарата актин-миозин для обеспечения успешной передачи.
Различия в полярности коллективно мигрирующих групп клеток обусловлены особенностями экспрессии поверхностных рецепторов, как хемокин CXCR4 и CXCR7 рецепторы в «лидерных» клетках. Факторы роста и хемокины, продуцируемые стромальными клетками, градиент диффузии обеспечивают внеклеточную индукцию поляризации клеток. Участие хемокинов, таких как SDF1 (CXCL12), фактор роста фибробластов (FGF) и трансформирующий фактор роста β (TGF-β), в этом процессе обсуждается.
Многое известно о участии TGF-β в канцерогенезе, при этом его роль двоякая. Тейлор и др. обратили на тот факт, что TGF-β, который действует в эпителиальных клетках молочной железы, как мощный опухолевый супрессор на ранних стадиях рака, может влиять на развитие опухоли через взаимодействие скогенными цитокинами. Повышенная экспрессия TGF-β связана с прогрессированием опухоли, что часто наблюдается, например, на более поздних стадиях рака груди. Роль TGF-β в эпителиально-стромальной миграции во время опухолевой прогрессии изучена недостаточно. Предполагается, что TGF-β является ключевым регулятором взаимодействий между опухолью и стромой, что способствует совместному развитию клеток при раке груди.
Было установлено, что лидерные клетки экспрессируют подопланин, трансмембранный гликопротеин, который экспрессируется в нормальных условиях в подоцитах почек, альвеолярных клетках легких 1 типа, клетках скелетных мышц, плаценте и т. д. Экспрессия подпланина в клетках рака молочной железы и инвазию клеток мигрени образования филоподий и одновременным сохранением экспрессии эгерина.
Были получены данные, указывающие на то, что коллективно мигрирующие раковые клетки могут использовать способность соседних мезенхимальных клеток восстановить матрикса и восстанавливать ее, и следовать по их «стопам». В экспериментах in vitro введение фибробластов в культуру вызывает коллективную миграцию опухолевых клеток в нижележащий матрикс в виде цепочек. Следовательно, фибробласты являются «проводником» для проникновения в опухолевые клетки, ремоделирования окружающего внеклеточного матрикса в пути с толстыми пучками коллагена по бокам и отсутствием матрикса в центре.
LIM-киназа, член группы одно из белковых семейств играет роль в развитии коллективной передачи опухолевых клеток. Известно, что этот участвует в регуляции развития инвадоподий, которые соответствуют структуре, типичные для злокачественных опухолевых клеток и ответственные за разрушение окружающей внеклеточной матрицы. Избыточная активация LIM-киназы проявляется при раке груди. Клетки опухоли груди с подавленной экспрессией гена LIM-киназы теряют способность к инвазии из-за потери способности разрушать внеклеточный матрикс.
Инвазия отдельных клеток или миграция отдельных клеток
Такой тип инвазивного роста, как одноклеточная инвазия, различается на основе обнаружения во время морфологического анализа отдельных опухолевых клеток, которые проникают в окружающие ткани независимо друг от друга. При этом типе опухолевой инвазии миграция одиночных клеток может происходить посредством двух различных типов поведения: мезенхимального и амебоидного. Ряд исследователей на возможность перехода от одного типа миграции к другому (от мезенхимального к амебоидному и наоборот, рис. 1) в случае инвазии единичных клеток. Эти переходы обычно проходят при изменении активности определенных клеточных молекул, когда опухолевые клетки должны адаптироваться к особенностям микроокружения.
Миграция мезенхимальных (фибробластоподобных) клеток
Мезенхимальные механизмы роста инвазивных клеток, напротив к амебоидному типу миграции, характеризуются протеканием более сложных процессов и необходимостью вовлечения в его большее количество клеточных мобильных клеток (рис.1).
Этот тип перемещения является типом из кератиноцитов в течение, гладкомышечных клеток и фибробластов. Мезенхимальные модели, теряют эпителиальную полярность и приобретают удлиненную форму веретена, напоминающую форму фибробластов, инвазия этого типа также называется «фибробластоподобной» миграцией. Мезенхимальная инвазия была обнаружена во время развития меланомы, фибросаркомы, глиобластомы и других злокачественных новообразований.
Большинство раковых клеток, которые отделяются из опухолевой массы и вторгаются в окружающие ткани, как известно, претерпевают определенные изменения, приобретая морфологические свойства и фенотип, характерные для мезенхимальных клеток. Эта трансформация злокачественной эпителиальной клетки, связанная с появлением в ней новых молекулярных и морфологических особенностей, получила название «эпителиально-мезенхимальный переход». Как уже упоминалось, это биологическое явление впервые было описано Э. Hay в 1995 году. Сегодня существование этого явления подтверждается результатами большого количества исследований, в которых изучались механизмы инвазии и метастазирования злокачественных опухолей. Мезенхимальный механизм инвазии считается следствием ЭМП, когда происходит активная дедифференцировка злокачественной эпителиальной опухоли, и многоклеточные группы начинают делиться на отдельные опухолевые клетки, приобретая мезенхимальный фенотип.
Ряд исследователей подчеркнули, что опухолевые клетки во время мезенхимального типа миграции проходят ряд конкретных последовательных шагов, которые составляют пятиступенчатую модель миграции. Этот цикл включает в себя следующие изменения: 1) образование выпячивания на одном из полюсов клетки - ламеллиподий или филоподий, возникающих в результате сокращений актинового цитоскелета под контролем малых GTPases Rac1 и Cdc42 с быстрым вовлечением интегринов семейства β1. ; 2) возникновение очаговой адгезии с участием интегринов β1 и β3 на участке контакта между внеклеточным матриксом и клеткой; 3) сборка фокальных контактов, которая основана на взаимодействиях, опосредованных интегрином, и активация протеолитических ферментов (матриксных металлопротеиназ, сериновых и треониновых протеаз, катепсинов) на границе раздела «клетка-матрица», что приводит к разрушению и ремоделированию окружающей среды. внеклеточный матрикс; 4) изменение поляризации актинового цитоскелета под контролем миозина II, возникновение сокращений клеточного тела; и 5) «вытягивание» задней кромки в сторону движения через вновь образованные дефекты в матричной структуре. Поскольку клетки, которые используют фибробластоподобный механизм инвазии, следуют описанным этапам миграции, их скорость движения невысока: около 0,1– мкм / мин.
Возможность протеолиза и ремоделирования тканевых структур объясняет этот факт что мезенхимальное движение опухолевой клетки сопровождается незначительными изменениями по сравнению с амебоидной миграцией в форме клетки и минимальной деформацией ядра. Несомненный интерес представляют результаты исследований, которые показывают, что поведение опухолевых клеток во время индивидуальной миграции зависит от жесткости окружающей матрицы. Например, мезенхимальная или протеолитическая модель миграции доминирует в условиях «жесткого» («плотного») окружающего матрикса. Высокая эффективность миграции отдельной клетки по мезенхимальному механизму в плотных тканях объясняется протеолизом за счет секреции различных протеаз и способностью образовывать очаговые контакты со стромальными элементами.
Поэтому стоит отметить что ключевыми моментами фибробластоподобного механизма инвазивного роста являются сильные силы адгезии на обоих полюсах клетки, а также между клетками и компонентами внеклеточного матрикса, выраженная экспрессия интегринов (семейства β1 и β3), протеолиз с разрушением и последующим ремоделированием тканей с образованием дефектов в матричной структуре и перемещением отдельной клетки или клеточных цепочек через дефекты. Деформация ядра минимальна, и наблюдается медленная скорость миграции клеток.
На основании подавления экспрессии соответствующих генов с использованием малых интерферирующих РНК, удельная активность GTPases Rac1 и Cdc42 была продемонстрирована как характерный признак мезенхимального типа инвазии. Подавление GTPase Rac1 через сигнальную активацию GTPase RhoA и ее эффектора, ROCK-киназы, приводит к блокированию мезенхимальной миграции опухолевых клеток.
Миграцияамебоидных клеток
Амебоидный механизм инвазивного роста - это наиболее эффективный способ передачи единичных опухолевых клеток. По всем своим характеристикам он похож на поведение и движение одноклеточного организма, такого как амеба Dictyostelium discoideum.
Использование антител, блокирующих интегрины или ингибиторы протеаз, в клинических испытаниях появления новых случаев появления новых вирусов в клетках с амебоидным типом миграции. Аналогичные результаты получены при изучении злокачественных опухолей in vivo. Установлена взаимосвязь между применением препаратов на основе ингибиторов матриксных металлопротеиназ в терапии рака и прогрессирования опухолевого процесса. Объяснение этой связи стало возможным только после опухолевых клеток, способных к амебоидной миграции. Эти, скорее всего, указывают на то, что в условиях снижения или полной утраты способности распространяться на окружающие ткани с помощью основных молекул, осуществляющих адгезию и разрушение внеклеточного матрикса, данные опухолевые клетки переходят к амебоидному механизму инвазии, который становится наиболее эффективным способом использования.
Этот тип транспорта был описан в циркулирующих стволовых клетках, лейкоцитах и некоторых типах опухолевых клеток. Согласно Zijl et al., Инвазивный рост амебоидного типа наблюдается при раке груди, лимфоме, мелкоклеточном раке легкого и раке простаты, а также меланоме.
В случае передачи амебоидных клеток, опухолевые клетки имеют установленное, что они имеют круглую или эллиптическую форму (рис. 1). Амебоидные клетки характеризуются быстрой деформируемостью, адаптацией своей формы к существующей структуре окружающего внеклеточного матрикса и проникновением через них через узкие пространства в сжатой форме. Движение и перемещение через последовательные высокоскоростные циклы расширения и сжатия клетки с расширением «пузырьковых» выступов клеточной мембраны. Эти пузыри позволяют клетке исследовать микросреду, чтобы найти наиболее подходящий маршрут движения, чтобы обойти препятствия, благодаря которым опухолевые клетки перемещаются через узкие промежутки во внеклеточном матриксе. Развивающиеся формы изменений клеток вызываются актиновым цитоскелетом коры головного мозга, который, в свою очередь, контролируется малой GTPase RhoA и ее эффектором, ROCK-киназой. Эта ГТФаза принадлежит к суперсемейству малых ГТФ-гидролаз, члены которых играют ключевую роль в инвазии амебоидного типа, поскольку они участвуют в передаче сигнала и таким образом, в регуляции широкого спектра процессов, происходящих в клетке, включая реорганизацию. актинового скелета во время перемещения цитоскелета.
Следует отметить, что миграция по амебоидному механизму инвазии сопровождается изменениями не только формы клетки, но и ядра, его ориентации и положения относительно к другим внутренним органеллам. Ядро, которое является наиболее прочным и жестким по сравнению с окружающей цитоскелет, органеллой, механически стабилизировано разветвленной сетью структурных белков, и поэтому его форма, скорее всего, часто не претевает значительных изменений. Однако амебоидный тип используется наиболее выраженной деформацией ядра, вызванной отсутствием протеолитической деградации окружающего матрикса. Коробка передач в удобном положении. Предполагается, что, подобно амебоидному движению лейкоцитов, ядро внутри одиночных мигрирующих опухолевых клеток продвигается к переднему краю.
В отличие от мезенхимального движения, амебоидная или непротеолитическая модель преобладает, когда окружающая матрица показывает относительно низкой жесткостью («мягкая» матрица). Например, амебоидная миграция опухолевых клеток в лимфатической и кровеносной системе как миграция в мягком матриксе.
Кондилис и Сегалл ли особенности особенности особенности линиям, MTC и MTLn3. в условиях in vitro и in vivo. Клетки MTLn3, обладающие высоким метастатическим потенциалом и мигрирующими, характеризуются более высоким уровнем экспрессии рецепторов эпидермального фактора роста (EGFR), чем клетки MTC с низким метастатическим потенциалом. Их миграция связана с наличием кровеносных сосудов и коллагенсодержащих волокон в окружающей матрице. Считается, что хемотаксис опухолевых клеток по направлению к кровеносным сосудам опосредуется сигнальными путями EGFR.
Амебоидный механизм инвазии имеет ряд отличительных особенностей. Для него характерно слабое взаимодействие между клетками и окружающей матрицей, а также отсутствие или слабые очаговые контакты. Отмечена возможность быстрой и нефокальной сборки рецепторов контактов с внеклеточным субстратом. Интегрины не важны при этом типе инвазивного роста. Важными аспектами являются отсутствие протеолиза в отношениях взаимодействия-матрица и отсутствие экспрессии протеолитических ферментов, разрушающих внеклеточный матрикс. Исследования in vitro показали, что в случае инвазивного роста амебоидного типа, вероятно, благодаря этим свойствам опухолевые клетки перемещаться с максимальной скоростью в культуре (20 мкм / мин).
Амебоидно-мезенхимальные и мезенхимально-амебоидные переходы
Мы уже отмечали наличие определенной степени пластичности и возможности «перехода» от одного типа передачи к другому (от мезенхимального типа к амебоидному). и наоборот) при инвазии отдельных клеток. Эти события, по-видимому, связаны с появлением изменений в настройках тканевого микроокружения (рис.1).
Эти изменения описываются как амебоидно-мезенхимальные и мезенхимально-амебоидные. переходы. Опухолевые клетки, использующие мезенхимальный тип передачи, могут определенным образом изменяться и переходить к амебоидному типу движения в условиях ослабления сигнальных и механических путей, которые непосредственно участвуют в стабилизации взаимодействия между структурами внеклеточного матрикса и злокачественными клетками. Однако данные были получены в основном экспериментальным путем. Описаны следующие механизмы, приводящие к переходу клеток от мезенхим к амебоидному типу инвазивного роста (мезенхимально-амебоидный переход): 1) снижение или полное прекращение перицеллюлярного протеолиза за счет применения ингибиторов протеаз; 2) снижение активности рецепторов интегрина и их взаимодействия с окружающими элементами стромы их антагонистами; 3) повышение и стабилизация активности малой ГТФазы RhoA и ее эффектора ROCK. Исследование, проведенное группой С. Бертона, дает интересный факт, указывает на то, что белок p27, несмотря на большое разнообразие функций, играет важную роль в контроле подвижности клеток. В частности, недостаток этой белка в условиях in vitro вызывает мезенхимально-амебоидный переход в клетках в трехмерном матриксе.
Некоторые авторы, изучающие механизмы инвазивного роста при индивидуальной передаче клеток, указывают на возможность амебоидного мезенхимального перехода, который происходит, обратным мезенхимальному переходу. Существует гипотеза, согласно которой основан механизм амебоидно-мезенхимального перехода, скорее всего, основан на той же молекулярной основе, и что единственный надежный процесс, определяющий возможность описанной трансформации, - это дисбаланс в активности членов семейства ГТФаз. и преобладание активности Rac над активностью RhoA. Механизмы, которые могли лежать в основе описанных изменений, остаются неясными.
Опухолевые клетки в пределах одной опухоли могут перемещаться одновременно как коллективно, так и индивидуально. В этом случае переход от индивидуальной передачи к коллективной является важным шагом на пути увеличения инвазивного и метастатического злокачественных новообразований. Например, клетки опухоли груди, отделившиеся от твердой массы, приобретают способность проникать в лимфатические сосуды. В настоящее время различают два механизма: эпителиально-мезенхимальный и коллективно-амебоидный переходы, с помощью которых образуются индивидуально мигрирующие опухолевые клетки (рис. 1). В свою очередь, последние, в особенности клетки, прошедшие ЭМП, способны в определенных условиях приобретать эпителиальный фенотип и образовывать опухолевые многоклеточные комплексы. Эта инверсия фенотипа получила название «мезенхимно-эпителиальный переход».
Эпителиально-мезенхимальный переход
В последнее время активно обсуждается эпителиально-мезенхимальный переходный механизм, в ходе которого опухолевые клетки отделяется от эпителиального слоя и приобретает подвижность (называемый рис. 1), так называемый «локомоторный» фенотип », который способствует инвазивному росту и метастазированию. Развитие процесса как ключевого фактора прогрессирования рака роста было показано in vitro с использованием определенных линий опухолей, а также экспериментальных моделей; однако установление развития ЭМП и их основных характеристик в условиях in vivo является сложной сложной.
ЭМП является причиной многих процессов морфогенеза. Считается, что в нормальных условиях (во время эмбриогенеза) ЭМП может индуцироваться HGF (фактор роста гепатоцитов), секретируемый фибробластами. HGF связывается со специфическими рецепторами c-Met, расположенными на мембранах эпителиальных клеток. Связывание с рецепторами активирует сигнальный путь с некоторыми белками системы малых ГТФаз (Cdc42, Rac, RhoA, RhoC), которые регулируют интенсивность полимеризации актиновых микрофиламентов и сократительную способность актин-миозиновых филаментов, что определяет интенсивность образования ламеллиподий и мощность прикрепленной к матрице клетки. В этом случае происходит значительная перестройка всего актин-миозинового цитоскелета и потеря межклеточных контактов Е-кадгерина. Во время канцерогенеза эпители подвергаются морфологической трансформации, которая фенотипически представляет собой ЕМТ, но происходит отсутствие соответствующей лиганда HGF. Эта трансформация в злокачественных опухолях может быть вызвана трансфекцией различных онкогенов. Во время трансформации опухолевые клетки могут покидать эпителиальный слой и двигаться подобно фибробластам, тем самым приобретая способность к инвазии и метастазированию.
Во время EMT происходят следующие основные события: злокачественные эпителиальные клетки теряют свою апикально-базальную полярность из-за нарушение плотных межклеточных контактов и потеря молекул клеточной адгезии (таких как E-кадгерин и интегрины); клеточный актиновый цитоскелет изменяется и подвергается ремоделированию стрессовых волокон, которые собираются в определенных клетках вблизи клеточной мембраны, где начинают формироваться специфические клеточные выступы; происходит разрушение базовой базальной мембраны эпителия, в результате чего опухолевые клетки, лишенные межклеточных контактов, становятся способными к инвазивному росту и проникновению в окружающий стромальный матрикс и начинают активную миграцию.
Было обнаружено, что ЭМП редко одинаково выражен во всей опухолевой ткани. Скорее всего, для этого процесса характерна разная интенсивность перехода клеток от эпителиального фена к мезенхимальному. В связи с этим некоторые исследователи описывают так называемую частичную ЭМП, в которой участвует большинство клеток инвазивного фронта (рис. 1). Частичная ЭМП - это состояние, когда клетки уже приобрели свойства, необходимые для успешной миграции, но продолжают сохранять межклеточные контакты. Этот фенотип был назван гибридным «эпителиально-мезенхимальным» фенотипом и был связан с особенностями, характерными для коллективно движущихся опухолевых клеток.
Taddei et al. Указали, что ЭМП развивается из-за индукции программ, связанных активацией ключевых факторов транскрипции, таких как TWIST1, Snail, Slug и ZEB1 / 2. Это приводит к нарушению прочных соединений кадгерина и активации миграции полярных клеток и протеолиза компонентов внеклеточного матрикса. различными секретируемыми протеазами, при этом функции рецепторов интегрина сохраняются. Экспериментально установлена роль фактора транскрипции Prrx1, который определяет способность клеток рака груди к инвазивному росту.
Было показано, что белки ZEB1 и ZEB2 с доменом цинкового пальца способны напрямую связываться с промоторами., тем самым индуцируя экспрессию генов мезенхимальных маркеров и подавляя экспрессию E-кадгерина и других эпителиальных маркеров.
Аналогичным образом Snail и Slug способны подавлять экспрессию гена E-кадгерина посредством прямого связывания с его промотора, а также продукции эпителиальных белков, таких как десмоплакин и клаудин, и активируют экспрессию виментина и матриксных металлопротеиназ, тем самым увеличивая миграцию клеток. Группа исследователей под руководством Санчеса-Тилло обнаружила, что фактор транскрипции Snail не встречается в нормальных эпителиальных клетках и что его обнаружение в клетках инвазивного фронта опухоли можно рассматривать как предиктор плохой выживаемости онкологических больных. Считается, что ZEB1 / 2, Snail и Slug индуцируются TGF-β, воспалительными цитокинами и гипоксией.
Коллективно-амебоидный переход
Основываясь на экспериментальных данных, ряд исследователей указывают на возможное существование так называемого коллективно-амебоидного перехода (рис. 1), когда опухолевые массы, вторгающиеся в окружающие ткани в виде коллективных многоклеточных групп, диссоциируют на отдельные мигрирующие клетки, которые используют амебоидное движение. Было показано, что это событие стало возможным благодаря применению ингибиторов интегриновых рецепторов семейства β1, поскольку эти молекулы играют ключевую роль как в формировании межклеточных контактов, так и во взаимодействиях между опухолевыми клетками и компонентами окружающей ткани. 25>
Мезенхимно-эпифизарный элиальный переход
На самом деле нет исследований, посвященных изучению механизмов, лежащих в основе мезенхимально-эпителиального перехода. Однако возможность такого явления признается. В этом случае говорят, что часто, например, при раке молочной железы и простая структура в отдаленных метастатических очагах ткани первичной опухоли. По мнению Фридла и Гилмора, на основании этих данных можно сделать несколько предположений. Во-первых, инвазия и метастазирование могут происходить без ЕМТ. Во-вторых, обнаружение единичных диссеминированных клеток при рутинном патологическом исследовании опухолевой ткани представляет собой довольно сложную информацию, а идентификация этих клеток при ЭМП практически невозможна. И, в-третьих, опухолевые клетки временно используют механизмы EMT для интравазации и распространения в отдаленные органы и ткани, где они возвращаются к эпителиальному фенотипу. Эта трансформация описывается как мезенхимально-эпителиальный переходный (MET). МЕТ индуцирован экспериментально, и индивидуально движущиеся клетки образ многоклеточные комплексы, но молекулярные механизмы МЕТ в физиологических условиях остаются неизвестными. Nguyen et al. обладает, что селективный ингибитор PD173074 рецептора 1 фактора роста фибробластов (FGFR1) ингибирует сигнальный путь MAPK, регулирующий активность белка AP-1, который, в свою очередь, индуцирует MET. Исследование возможности использования ингибитора PD173074 в качестве лекарственного средства, проведенное на определенных линиях опухолевых клеток, выявило отчетливое подавление роста опухоли, миграционной способности и инвазии. В этом случае наблюдалось снижение экспрессии генов Улитка и матриксной металлопротеиназы 3, 10, 12 и 13 и повышение экспрессии гена E-кадгерина.
На протяжении многих лет наша исследовательская группа изучала особенности прогресса в зависимости от внутриопухолевой гетерогенности. Особое внимание было уделено фенотипическому разнообразию первичной опухоли при инвазивной карциноме без особого типа, которая составляет основную часть (80%) всех гистологических типов рака груди.
Несмотря на значительное структурное разнообразие опухолей. При первичной молочной железы можно выделить пять основных типов морфологических структур: альвеолярные, трабекулярные, тубулярные и солидные структуры и дискретные группы опухолевых клеток (рис. 2). Альвеолярные структуры соответствуют скопления опухолевых клеток круглой или неправильной формы. Морфология клеток, образующихся типичных клеток с умеренной цитоплазмой и округлыми ядрами клеток с гиперхроматическими ядрами неправильной и умеренной цитоплазмой формы. Трабекулярные структуры представляют собой либо короткие линейные ассоциации, образованные одним коротким, довольно мономорфным клеткам, либо широкие кластеры клеток, состоящие из двух рядов клеток среднего размера с умеренной цитоплазмой и круглыми нормохромными или гиперхроматическими ядрами. Трубчатые структуры образованы одним или двумя рядами довольно мономорфных клеток с округлыми нормохромными ядрами. Твердые структуры представляют собой поля различных размеров и форм, состоящих из мелких клеток с умеренной цитоплазмой и мономорфными ядрами, либо из крупных клеток с обильной цитоплазмой и с помощью обильных цитоплазмов. Дискретные группы клеток встречаются в виде кластеров от одной до четырех клеток с различной морфологией.
 Внутриопухолевая морфологическая неоднородность при инвазивной карциноме молочной железы. Показано разнообразие инвазивного роста молочной железы, которое можно разделить на пять основных морфологических структур: альвеолярные (Alv), трабекулярные (Trab), тубулярные (ванна), твердые (твердые) структуры и дискретные группы опухолевых клеток (Discr).. Окрашивание гематоксилином и эозином. Увеличение 200x.
Внутриопухолевая морфологическая неоднородность при инвазивной карциноме молочной железы. Показано разнообразие инвазивного роста молочной железы, которое можно разделить на пять основных морфологических структур: альвеолярные (Alv), трабекулярные (Trab), тубулярные (ванна), твердые (твердые) структуры и дискретные группы опухолевых клеток (Discr).. Окрашивание гематоксилином и эозином. Увеличение 200x. По данным, накопленным на сегодняшний день, можно предположить, что разные морфологические структуры соответствуют типам инвазии. Таким образом, альвеолярные, трабекулярные и твердые структуры, для которых характерно межклеточных контактов, можно отнести к морфологическим проявлениям коллективной передачи, а дискретные группы опухолевых клеток - к проявлениям индивидуальной передачи. Интересно, что первая серия данных, полученное при исследовании экспрессии генов клет-адгезии, полностью подтверждает эту гипотезу. Например, снижение активности генов кадгеринов, отвечающих за межклеточные контакты, в следующем порядке: солидные - альвеолярные и трабекулярные структуры - дискретные группы опухолевых клеток. В этом случае количество экспрессируемых генов интегринов, участвующих в адгезии опухолевых клеток к внеклеточному матриксу, уменьшилось в следующем порядке: солидные и альвеолярные - трабекулярные структуры - дискретные группы опухолевых клеток.
Инвазивный рост и развитие лекарственной устойчивости - связанные процессы, которые стимулируют роль в прогрессировании опухоли, в частности, в метастазировании. Весьма вероятно, что одни и те же сигнальные пути участвуют в передаче клеток и развитии устойчивости опухоли к терапии.
Мигрирующие опухолевые клетки (независимо от типа) более устойчивы к химиотерапии и лучевой терапии, чем неподвижные клетки. Во многом это связано с тем, что мигрирующие клетки временно теряют способность делиться. Это также факт, что движущиеся опухолевые клетки проявляют повышенную активность антиапоптотических генов, что вызывает устойчивость к химиотерапевтическим препаратам, направленным на индукцию запрограммированной гибели клеток. Кроме того, известно, что клетки в состоянии EMT также проявляют химиорезистентность. Эта лекарственная устойчивость из-за индукции во время ЕМТ-синтеза белков семейства ABC, ответственных за отток химиотерапевтических препаратов из клеток. Основные факторы транскрипции, которые запускают EMT и в то же время положительно регулируют активность переносчиков ABC, включая TWIST1, Snail и т. Д.
Недавно полученные данные указывают на сильную связь между коллективной миграцией и устойчивостью к лучевой и химиотерапии. Согласно нашим собственным исследованиям, опухоли молочной железы, устойчивые как альвеолярные, так и трабекулярные структуры, а также демонстрирующие значительное морфологическое разнообразие, характеризуются повышенной лекарственной устойчивостью. Интересно, что вклад трабекулярных структур в химиорезистентность, вероятно, объясняется высокой активностью транспортеров ABC в опухолевых клетках того или иного морфологического варианта. Напротив, резистентность опухолей груди, альвеолярная устойчивость, объясняется другими, но не установленными причинами.
Инвазивный рост и его фенотипическое разнообразие связаны как напрямую, так и через развитие лекарственной устойчивости с метастазами. Циркулирующие опухолевые клетки, которые вызывают развитие будущих метастазов, вызывают развитие инвазии и последующего проникновения опухолевых клеток в лимфатические или кровеносные сосуды. Не только одиночные мигрирующие опухолевые клетки, но и группы клеток обладают способностью к интравазации. Есть предположение, что коллективная миграция чаще приводит к метастазированию, чем индивидуальная миграция. Новаторские исследования на животных в моделях показывают, что метастазы чаще наблюдаются после инъекции опухолевых кластеров, а не отдельных опухолевых клеток. Кроме того, циркулирующие скопления опухолевых клеток были обнаружены в крови пациентов с другими видами рака. Предполагалось, что коллективная интравазация связана с VEGF-зависимым формированием расширенной сосудистой сети и накоплением внутривазированных опухолевых кластеров. Кроме того, опухолевые клетки могут поступать в кровоток через поврежденные сосуды или за счет взаимодействия с клетками в состоянии EMT и связанными с раком фибробластами, которые разрушают внеклеточный матрикс протеазами. Зость метастазов от коллективной зависимости от исходных данных сохраняется результаты собственных исследований. Например, у пациентов с высокой скоростью лимфогенных метастазов увеличиваются масштабы роста этого типа у женщин в пременопаузе с увеличенным количеством различных типов морфологических структур. Последняя также носит количественный характер: лимфогенные метастазы выявляются при большом количестве альвеолярных структур в опухолях молочной железы. Кроме того, пациенты с альвеолярными структурами в опухолях имели низкую выживаемость без метастазов (наши собственные неопубликованные данные).
Установленная взаимосвязь между альвеолярными структурами, как одно из проявлений коллективной передачи, скорость лимфогенного и гематогенного метастаз позволяет сделать следующие предположения. По-видимому, клеточные элементы альвеолярных структур отличаются набором биологических свойств, определяющих метастатический фенотип. Более четкая взаимосвязь между альвеолярными структурами и лимфогенными метастазами в период менопаузы предполагает определенную роль эстрогенов, в том числе их производство in situ, что опухолевые клетки альвеолярных структур приобретают метастатический фенотип через лимфогенный путь.
. контроля ». »Прогрессирования заболеваний.
По степени инвазии рак может быть классифицирован как in situ, когда злокачественные клетки присутствуют в виде опухоли, но не метастазируют и не проникают за пределы слоя или типа ткани, где она возникла. Например, рак эпителиального происхождения с такими признаками называется карциномой in situ и определяется как рак, не проникший за пределы базальной мембраны. Напротив, инвазивная карцинома проникла за пределы базальной мембраны.
Не вызывает сомнения значимость исследований морфологических проявлений и молекулярно-генетических механизмов инвазии и метастазирования злокачественных опухолей. Результаты исследований показывают, что миграция опухолевых клеток при инвазивном росте может происходить как через отдельные клетки, так и через группы клеток. Такое разнообразие типов клеток, вероятно, приводит к развитию внутриопухолевой гетерогенности, которая представлена, например, при раке молочной железы - разными морфологическими структурами: альвеолярными, трабековыми, солидными структурами и отдельными группами опухолевых клеток. Известен биохимических и молекулярно-генетических механизмов, которые позволяют злокачественным клеткам проникать в окружающие ткани и приобретать способность распространяться далеко за пределы первичного очага опухоли, вызывая вторичные метастатические очагов в отдаленных органах и тканях. Однако, несмотря на достигнутый прогресс, неисследованные вопросы, касающиеся возможной взаимосвязи между различными типами инвазивного роста клеток и функций гематогенного метастазирования, особенностями прогрессирования заболеваний, а также эффективностью выбранной терапии. Решение этих проблем может оказаться большим подспорьем в прогнозировании заболеваний и, возможно, в разработке новых подходов к ведению онкологических больных.
Эпителиально-мезенхимальный переход EMT MET мезенхимально-эпителиальный переход ГТФазы гуанозинтрифосфатазы
.