
Войти
| Иммуноглобулиноподобный рецептор | |
|---|---|
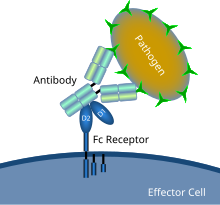 Схематическая диаграмма, показывающая взаимодействие рецептора Fc с микробным патогеном, покрытым антителами Схематическая диаграмма, показывающая взаимодействие рецептора Fc с микробным патогеном, покрытым антителами | |
| Идентификаторы | |
| Символ | рецептор Fc |
| Мембранома | 10 |
A рецептор Fc представляет собой белок, обнаруживаемый на поверхности определенных клеток, включая, среди прочего, B-лимфоциты, фолликулярные дендритные клетки, естественные киллерные клетки, макрофаги, нейтрофилы, эозинофилы, базофилы, человеческие тромбоциты и тучные клетки - которые вносят вклад в защитные функции иммунной системы. Его название происходит от его специфичности связывания с частью антитела, известной как область Fc (фрагмент, кристаллизующийся). Рецепторы Fc связываются с антителами, которые прикреплены к инфицированным клеткам или вторгаются патогенам. Их активность стимулирует фагоцитарные или цитотоксические клетки для уничтожения микробов, или инфицированные клетки посредством опосредованного антителами фагоцитоза или антителозависимого клеточно-опосредованная цитотоксичность. Некоторые вирусы, такие как флавивирусы, используют рецепторы Fc, чтобы помочь им инфицировать клетки с помощью механизма, известного как антитело-зависимое усиление инфекции.
Существует несколько различных типов рецепторов Fc (сокращенно FcR), которые классифицируются на основе типа антитело, которое они распознают. Латинская буква, используемая для идентификации типа антитела, преобразуется в соответствующую греческую букву, которая помещается после части имени «Fc». Например, те, которые связываются с наиболее распространенным классом антител, IgG, называются рецепторами Fc-гамма (FcγR), те, которые связывают IgA, называются рецепторами Fc-альфа (FcαR). и те, которые связывают IgE, называются рецепторами Fc-эпсилон (FcεR). Классы FcR также различаются по экспрессирующим их клеткам (макрофаги, гранулоциты, естественные клетки-киллеры, Т- и В-клетки) и сигнальным свойствам каждого рецептора.
Все рецепторы Fcγ (FcγR) принадлежат к суперсемейству иммуноглобулинов и являются наиболее важными рецепторами Fc для индукции фагоцитоза опсонизированных (маркированных) микробов. Это семейство включает несколько членов, FcγRI (CD64), FcγRIIA (CD32 ), FcγRIIB (CD32), FcγRIIIA (CD16a), FcγRIIIB (CD16b), которые различаются по своей аффинности антител. к их различной молекулярной структуре. Например, FcγRI связывается с IgG сильнее, чем FcγRII или FcγRIII. FcγRI также имеет внеклеточную часть, состоящую из трех иммуноглобулиновых (Ig) -подобных доменов, что на один домен больше, чем у FcγRII или FcγRIII. Это свойство позволяет FcγRI связывать единственную молекулу IgG (или мономер ), но все рецепторы Fcγ должны связывать несколько молекул IgG в пределах иммунного комплекса для активации.
Рецепторы Fc-гамма различаются по своему сродству к IgG, и аналогично разные подклассы IgG обладают уникальным сродством к каждому из рецепторов Fc гамма. Эти взаимодействия дополнительно регулируются гликаном (олигосахаридом) в положении CH2-84.4 IgG. Например, создавая стерические препятствия, фукоза, содержащая гликаны CH2-84.4, снижает сродство IgG к FcγRIIIA. Напротив, гликаны G0, в которых отсутствует галактоза и заканчиваются вместо этого фрагментами GlcNAc, имеют повышенное сродство к FcγRIIIA.
Другой FcR экспрессируется на нескольких типах клеток и по структуре сходен с MHC класса I. Этот рецептор также связывает IgG и участвует в сохранении этого антитела. Однако, поскольку этот рецептор Fc также участвует в передаче IgG от матери либо через плаценту к ее плоду, либо с молоком грудному ребенку , он называется неонатальным рецептором Fc (FcRn ). Недавние исследования показали, что этот рецептор играет роль в гомеостазе сывороточных уровней IgG.
Только один рецептор Fc принадлежит к подгруппе FcαR, которая называется FcαRI (или CD89). FcαRI обнаружен на поверхности нейтрофилов, эозинофилов, моноцитов, некоторых макрофагов (включая клетки Купфера ) и некоторых дендритных клеток. Он состоит из двух внеклеточных Ig-подобных доменов и является членом суперсемейства иммуноглобулинов и семейства рецепторов многоцепочечного иммунного распознавания (MIRR). Он сигнализирует, связываясь с двумя сигнальными цепями FcRγ. Другой рецептор также может связывать IgA, хотя он имеет более высокое сродство к другому антителу, называемому IgM. Этот рецептор называется рецептором Fc-alpha / mu (Fcα / μR) и представляет собой трансмембранный белок типа I. Имея один Ig-подобный домен во внеклеточной части, этот рецептор Fc также является членом суперсемейства иммуноглобулинов.
Известны два типа FcεR:
| Название рецептора | лиганд основного антитела | Аффинность к лиганду | Распределение клеток | Эффект после связывания с антителом |
| FcγRI (CD64 ) | IgG1 и IgG3 | High (Kd ~ 10 M) | Макрофаги. Нейтрофилы. Эозинофилы. Дендритные клетки | Фагоцитоз. Активация клеток. Активация респираторного взрыва. Индукция уничтожения микробов |
| FcγRIIA (CD32 ) | IgG | Низкая (Kd>10 M) | Макрофаги. Нейтрофилы. Эозинофилы. Тромбоциты. Клетки Лангерганса | Фагоцитоз. Дегрануляция (эозинофилы) |
| FcγRIIB1 (CD32) | IgG | Низкий (Kd>10 M) | В-клетки. Тучные клетки | Нет фагоцитоза. Ингибирование клеточной активности |
| FcγRIIB2 (CD32) | IgG | Низкий (Kd>10 M) | Макрофаги. Нейтрофилы. Эозинофилы | Фагоцитоз. Ингибирование це Активность II |
| FcγRIIIA (CD16a ) | IgG | Низкая (Kd>10 M) | NK-клетки. Макрофаги (определенные ткани) | Индукция антителозависимой клеточно-опосредованной цитотоксичности (ADCC). Индукция высвобождения цитокинов макрофагами |
| FcγRIIIB (CD16b ) | IgG | Низкая (Kd>10 M) | эозинофилы. макрофаги. нейтрофилы. тучные клетки. фолликулярные дендритные клетки | индукция уничтожения микробов |
| FcεRI | IgE | Высокая (Kd ~ 10 M) | Тучные клетки. Эозинофилы. Базофилы. Клетки Лангерганса. Моноциты | Дегрануляция. Фагоцитоз |
| FcεRII (CD23 ) | IgE | Низкий (Kd>10 M) | B-клетки. Эозинофилы. клетки Лангерганса | Возможная молекула адгезии. Транспорт IgE через эпителий кишечника человека. Механизм положительной обратной связи для усиления аллергической сенсибилизации (В-клетки) |
| FcαRI (CD89 ) | IgA | Низкий (Kd>10 M) | Моноциты. Макрофаги. Neu трофилы. Эозинофилы | Фагоцитоз. Индукция уничтожения микробов |
| Fcα / μR | IgA и IgM | Высокая для IgM, Средняя для IgA | В-клетки. мезангиальные клетки. макрофаги | эндоцитоз. индукция уничтожения микробов |
| FcRn | IgG | моноциты. макрофаги. Дендритные клетки. Эпителиальные клетки. Эндотелиальные клетки. Гепатоциты. | Переносят IgG от матери к плоду через плаценту. Переносят IgG от матери к ребенку в молоке. Защищают IgG от деградация |
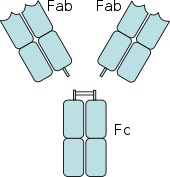 Антитело имеет области Fab (фрагмент, связывающий антиген) и Fc (фрагмент, кристаллизующийся). Рецепторы Fc связываются с областью Fc.
Антитело имеет области Fab (фрагмент, связывающий антиген) и Fc (фрагмент, кристаллизующийся). Рецепторы Fc связываются с областью Fc. Рецепторы Fc обнаружены на ряде клеток иммунной системы, включая фагоциты, такие как макрофаги и моноциты, гранулоциты, такие как нейтрофилы и эозинофилы, и лимфоциты врожденной иммунной системы (естественные клетки-киллеры ) или адаптивной иммунной системой (например, B-клетки ). Они позволяют этим клеткам связываться с антителами, которые прикреплены к поверхности микробов или инфицированных микробами клеток, помогая этим клеткам идентифицировать и устранять микробные патогены. Рецепторы Fc связывают антитела в своей области Fc (или хвосте), взаимодействие, которое активирует клетку, которая обладает рецептором Fc. Активация фагоцитов - наиболее частая функция, приписываемая рецепторам Fc. Например, макрофаги начинают поглощать и убивать покрытый IgG патоген посредством фагоцитоза после задействования их рецепторов Fcγ. Другой процесс с участием Fc-рецепторов называется зависимой от антител клеточной цитотоксичностью (ADCC). Во время ADCC рецепторы FcγRIII на поверхности естественных клеток-киллеров (NK) стимулируют NK-клетки высвобождать цитотоксические молекулы из их гранул, чтобы убить покрытые антителами клетки-мишени. FcεRI выполняет другую функцию. FcεRI представляет собой рецептор Fc на гранулоцитах, который участвует в аллергических реакциях и защите от паразитарных инфекций. Когда присутствует соответствующий аллергический антиген или паразит, перекрестное сшивание по крайней мере двух молекул IgE и их рецепторов Fc на поверхности гранулоцита заставляет клетку быстро высвобождать предварительно сформированные медиаторы из своих гранул. 231>
Fc-гамма-рецепторы относятся к группе некаталитических тирозин-фосфорилированных рецепторов, которые имеют сходную передачу сигналов путь, включающий фосфорилирование остатков тирозина. Рецепторы генерируют сигналы в своих клетках через важный мотив активации, известный как мотив активации тирозина иммунорецептора (ITAM). ITAM представляет собой специфическую последовательность аминокислот (YXXL), дважды встречающуюся в тесной последовательности во внутриклеточном хвосте рецептора. Когда фосфатные группы добавляются к тирозиновому (Y) остатку ITAM закрепленными на мембране ферментами из семейства Src-киназ, в ячейке генерируется сигнальный каскад . Эта реакция фосфорилирования обычно следует за взаимодействием рецептора Fc с его лигандом. ITAM присутствует во внутриклеточном хвосте FcγRIIA, и его фосфорилирование вызывает фагоцитоз в макрофагах. FcγRI и FcγRIIIA не имеют ITAM, но могут передавать активирующий сигнал своим фагоцитам, взаимодействуя с другим белком, который имеет. Этот адаптерный белок называется субъединицей Fcγ и, как и FcγRIIA, содержит две последовательности YXXL, которые характерны для ITAM.
Присутствия только одного мотива YXXL недостаточно для активации клеток, и он представляет собой мотив (I / VXXYXXL), известный как ингибиторный мотив на основе тирозина иммунорецептора (ITIM). FcγRIIB1 и FcγRIIB2 имеют последовательность ITIM и являются ингибирующими рецепторами Fc; они не вызывают фагоцитоз. Ингибирующее действие этих рецепторов контролируется ферментами, удаляющими фосфатные группы из остатков тирозина; фосфатазы SHP-1 и SHIP-1 ингибируют передачу сигналов рецепторами Fcγ. Связывание лиганда с FcγRIIB приводит к фосфорилированию тирозина мотива ITAM. Эта модификация генерирует сайт связывания для фосфатазы, домен узнавания SH2. Отмена передачи сигналов активации ITAM вызывается ингибированием протеинтирозинкиназ семейства Src и гидролизом мембраны PIP3, прерывая дальнейшую передачу сигналов ниже по течению активирующими рецепторами, такими как активация FcγRs, TCR, BCR и рецепторов цитокинов (например, c-Kit
Отрицательная передача сигналов с помощью FcγRIIB в основном важна для регуляции активированных В-клеток. Положительная передача сигналов В-клеток инициируется связыванием чужеродного антигена с поверхностным иммуноглобулином. Выделяется одно и то же антиген-специфическое антитело, которое может подавлять обратную связь или способствовать передаче негативных сигналов. Этот негативный сигнал обеспечивается FcγRIIB: эксперименты с использованием мутантов с делецией В-клеток и доминантно-отрицательных ферментов твердо установили важную роль SH2-домен-содержащей инозитол-5-фосфатазы (SHIP) в отрицательной передаче сигналов. Отрицательная передача сигналов через SHIP, по-видимому, ингибирует путь Ras через конкуренцию домена SH2 с Grb2 и Shc и может включать потребление внутриклеточных липидных медиаторов, которые действуют как активаторы аллостерических ферментов или способствуют проникновению внеклеточного Ca2 +.
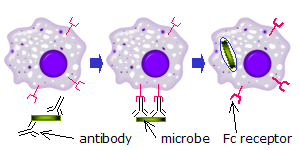 Fc-рецепторы распознают микробы, которые были связаны антителами. Взаимодействие между связанными антителами и рецептором Fc клеточной поверхности активирует иммунную клетку, чтобы убить микроб. В этом примере показан фагоцитоз опсонизированного микроба.
Fc-рецепторы распознают микробы, которые были связаны антителами. Взаимодействие между связанными антителами и рецептором Fc клеточной поверхности активирует иммунную клетку, чтобы убить микроб. В этом примере показан фагоцитоз опсонизированного микроба. Когда молекулы IgG, специфичные для определенного антигена или поверхности компонента, связываются с патогеном с их Fab-областью (фрагмент антигенсвязывающей области), их Fc-области направлены наружу, в непосредственной близости от фагоцитов. Фагоциты связывают эти области Fc своими рецепторами Fc. Между рецептором и антителом образуется множество низкоаффинных взаимодействий, которые вместе прочно связывают микроб, покрытый антителами. Низкая индивидуальная аффинность не позволяет рецепторам Fc связывать антитела в отсутствие антигена и, следовательно, снижает вероятность активации иммунных клеток в отсутствие инфекции. Это также предотвращает агглютинацию (свертывание) фагоцитов антителом, когда антиген отсутствует. После связывания патогена взаимодействия между Fc-областью антитела и Fc-рецепторами фагоцита приводят к инициации фагоцитоза. Патоген поглощается фагоцитом в результате активного процесса, включающего связывание и высвобождение комплекса Fc-область / Fc-рецептор, пока клеточная мембрана фагоцита полностью не закроет патоген.
Рецептор Fc на NK-клетках распознает IgG, который связан с поверхностью инфицированной патогеном клетки-мишени и называется CD16 или FcγRIII. Активация FcγRIII IgG вызывает высвобождение цитокинов, таких как IFN-γ, которые передают сигнал другим иммунным клеткам, и цитотоксических медиаторов, таких как перфорин и гранзим., которые проникают в клетку-мишень и способствуют гибели клеток, вызывая апоптоз. Этот процесс известен как антителозависимая клеточно-опосредованная цитотоксичность (ADCC). FcγRIII на NK-клетках также может связываться с мономерным IgG (то есть с IgG, который не связан с антигеном). Когда это происходит, рецептор Fc ингибирует активность NK-клетки.
 Активация дегрануляции тучных клеток посредством взаимодействия IgE с FcεRI. 1 = антиген; 2 = IgE; 3 = FcεRI; 4 = предварительно сформированные медиаторы (гистамин, протеазы, хемокины, гепарин); 5 = гранулы; 6 - тучная клетка; 7 - новообразованные медиаторы (простагландины, лейкотриены, тромбоксаны, фактор активации тромбоцитов)
Активация дегрануляции тучных клеток посредством взаимодействия IgE с FcεRI. 1 = антиген; 2 = IgE; 3 = FcεRI; 4 = предварительно сформированные медиаторы (гистамин, протеазы, хемокины, гепарин); 5 = гранулы; 6 - тучная клетка; 7 - новообразованные медиаторы (простагландины, лейкотриены, тромбоксаны, фактор активации тромбоцитов) антитела IgE связываются с антигенами из аллергенов. Эти связанные с аллергеном молекулы IgE взаимодействуют с рецепторами Fcε на поверхности тучных клеток. Активация тучных клеток после вовлечения FcεRI приводит к процессу, называемому дегрануляцией, в результате чего тучные клетки высвобождают предварительно сформированные молекулы из своих цитоплазматических гранул; они представляют собой смесь соединений, включающих гистамин, протеогликаны и сериновые протеазы. Активированные тучные клетки также синтезируют и секретируют медиаторы, производные липидов (такие как простагландины, лейкотриены и фактор активации тромбоцитов ) и цитокины. (например, интерлейкин 1, интерлейкин 3, интерлейкин 4, интерлейкин 5, интерлейкин 6, интерлейкин 13, фактор некроза опухоли альфа, GM-CSF и несколько хемокинов. Эти медиаторы способствуют воспалению за счет привлечения других лейкоцитов.
Крупные паразиты, такие как гельминт (червь) Schistosoma mansoni, слишком велики для поглощения фагоцитами. Они также имеют внешнюю структуру, называемую покровом, которая устойчива к атакам веществ, выделяемых макрофагами и тучными клетками. Однако эти паразиты могут покрываться IgE и распознаваться FcεRI на поверхности эозинофилов. Активированные эозинофилы высвобождают преформированные медиаторы, такие как как основной основной белок и ферменты, такие как пероксидаза, против которых гельминты не устойчивы. Взаимодействие рецептора FcεRI с частью Fc связанного с гельминтом IgE заставляет эозинофил высвобождать эти молекулы по механизму, аналогичному механизму NK-клетки во время ADCC.
CD4 + Т-клетки (зрелые Т h клетки ) помогают В-клеткам, вырабатывающим антитела. При патологии заболевания наблюдается несколько подмножеств активированных эффекторных CD4 + Т-клеток. Более ранние исследования, обобщенные Сандерсом и Линчем в 1993 г., предполагали критическую роль FcR в иммунных ответах, опосредованных CD4 + Т-клетками, и предполагали образование совместного сигнального комплекса между FcR и TCR на поверхности клетки. Chauhan и соавторы сообщили о совместной локализации меченых ICs с комплексом CD3 на активированной поверхности CD4 + Т-клеток, что, таким образом, предполагает сосуществование FcR вместе с комплексом TCR. Оба этих рецептора образуют апикальную структуру на мембране активированных CD4 + Т-клеток, что свидетельствует о боковом движении этих рецепторов. Совместная миграция FcR с TCR и комплексом BCR наблюдается на поверхности клеток, и цитоконъюгаты T: B-клеток демонстрируют это сосуществование в точке контакта. Предыдущий обзор показал, что экспрессия FcR на CD4 + Т-клетках остается открытым вопросом. Это установило текущую парадигму, согласно которой Т-клетки не экспрессируют FcR, и эти результаты никогда не подвергались сомнению и экспериментально не проверялись. Чаухан и его коллеги показали связывание иммунных комплексов (IC), лиганда FcR, с активированными CD4 + Т-клетками. Экспрессия CD16a индуцируется в активированных наивных CD4 + Т-клетках человека, которые экспрессируют CD25, CD69 и CD98, и лигирование с IC приводит к образованию эффекторных клеток памяти. Передача сигнала CD16a опосредуется фосфорилированием Syk (pSyk).
В настоящее время исследование предполагает индуцированную экспрессию CD32a при активации CD4 + Т-клеток человека, аналогичную CD16a. Экспрессия CD32a на CD4 + Т-клетках также была подтверждена тремя независимыми исследованиями исследователей ВИЧ-1. Экспрессия CD16a и CD32a в подгруппе активированных CD4 + Т-клеток теперь подтверждена. FcR на поверхности клетки при связывании с IC, состоящими из нуклеиновых кислот, запускают продукцию цитокинов и активируют пути восприятия нуклеиновых кислот. FcR присутствуют как на поверхности клетки, так и в цитозоле. Передача сигнала CD16a усиливает экспрессию толл-подобных рецепторов, чувствительных к нуклеиновым кислотам, и перемещает их на поверхность клетки. CD16a - это новый костимулирующий сигнал для CD4 + Т-клеток человека, который успешно заменяет потребность в CD28 во время аутоиммунитета. На аутоиммунном фоне CD4 + Т-клетки обходят требование косигнализации CD28 для полной активации. Кроме того, блокада косигнализации CD28 не подавляет развитие клеток TFH, ключевой субпопуляции для генерации аутоантител, продуцирующих аутореактивные В-клетки плазмы. Для иммунного гомеостаза необходим баланс между костимулирующими и тормозными сигналами. Чрезмерная костимуляция и / или недостаточное совместное ингибирование приводит к нарушению толерантности и аутоиммунитету. Опосредованная CD16a костимуляция обеспечивает положительный сигнал в активированных CD4 + Т-клетках, а не в покоящихся клетках, в которых отсутствует экспрессия FcγR.