
Войти
Генетическая кластеризация человека - это степень, в которой генетическая изменчивость человека может быть разделена на небольшое количество групп или кластеров. Ведущий метод анализа использует математический кластерный анализ степени сходства генетических данных между людьми и группами, чтобы сделать выводы о популяционных структурах и отнести людей к предполагаемым предковым группам. Аналогичный анализ можно провести с помощью анализа главных компонентов, и в нескольких недавних исследованиях используются оба метода.
Анализ генетической кластеризации изучает степень, в которой региональные группы отличаются генетически, категоризацию индивидов в кластеры и что можно узнать о происхождении человека из этих данных. Существует широкое научное согласие с тем, что относительно небольшая часть генетических вариаций человека происходит между популяциями, континентами или кластерами. Однако исследователи генетической кластеризации расходятся во мнениях относительно того, является ли генетическая изменчивость в основном клинальной или кластеры, выведенные математически, важны и полезны с научной точки зрения.
Один из основных вопросов, касающихся распределения генетического разнообразия человека, связан со степенью общих генов между наблюдаемыми кластерами. Неоднократно наблюдалось, что большая часть изменений, наблюдаемых в глобальной человеческой популяции, обнаруживается внутри популяций. Этот вариант, как правило, рассчитывается с использованием Sewall Wright «S индекса фиксации (F ST), который является оценкой между изменением в пределах группы. Степень генетической изменчивости человека немного отличается в зависимости от изучаемого типа гена, но в целом принято утверждать, что ~ 85% генетической изменчивости обнаруживается внутри групп, ~ 6-10% между группами на одном континенте и ~ 6–10% находится между континентальными группами. Райан Браун и Джордж Армелагос описали это как «множество исследований [которые] пришли к выводу, что схемы расовой классификации могут составлять лишь незначительную часть генетического разнообразия человека», включая исследования, перечисленные в таблице ниже.
| Авторы) | Год | заглавие | Характеристика изучена | Доля вариаций внутри групп (а не среди населения) |
|---|---|---|---|---|
| Левонтин | 1972 г. | Распределение человека разнообразие | 17 групп крови | 85,4% |
| Barbujani et al. | 1997 г. | Распределение разнообразия ДНК человека | 79 ПДРФ, 30 микросателлитных локусов | 84,5% |
| Зайльстад, Минч и Кавалли-Сфорца | 1998 г. | Генетические данные о более высокой скорости миграции женщин среди людей | 29 аутосомных микросателлитных локусов | 97,8% |
| 10 Y-хромосома микросателлитные локусы | 83,5% |
Эти средние числа, однако, не означают, что каждая популяция обладает равным разнообразием. В самом деле, некоторые популяции человека содержат гораздо больше генетическое разнообразие, чем другие, что согласуется с вероятным африканским происхождением из современных людей. Следовательно, популяции за пределами Африки могут подвергаться серийным эффектам основателей, которые ограничивают их генетическое разнообразие.
Статистика F ST подверглась критике со стороны AWF Эдвардса, Джеффри Лонга и Рика Киттлса. Британский статистик и биолог-эволюционист AWF Эдвардс осудил методологию Левонтина за то, что он основывал свои выводы на простом сравнении генов, а скорее на более сложной структуре частот генов. Возражение Лонга и Киттлса также носит методологический характер: по их мнению, F ST основан на ошибочном исходном предположении, что все популяции содержат одинаково генетически разнообразных членов и что континентальные группы разошлись одновременно. Сарич и Миле также утверждали, что оценки генетических различий между людьми из разных популяций занижают различия между группами, потому что они не принимают во внимание диплоидность человека.
Кейт Ханли, Грасиела Кабана и Джеффри Лонг создали пересмотренную статистическую модель для учета неравно расходящихся популяционных линий и местных популяций с разной степенью разнообразия. В их статье 2015 года эта модель применяется к выборке проекта Human Genome Diversity Project из 1037 человек в 52 популяциях. Они обнаружили, что наименее разнообразная исследованная популяция, суруи, «укрывает почти 60% всего разнообразия видов». Лонг и Киттлс ранее отметили, что африканцы сокото обладают практически всем генетическим разнообразием человека. Их анализ также показал, что неафриканские популяции представляют собой таксономическую подгруппу африканских популяций, что «некоторые африканские популяции в равной степени связаны с другими африканскими популяциями и неафриканскими популяциями», и что «за пределами Африки региональные группы популяций вложены друг в друга. внутри друг друга, и многие из них не являются монофилетическими ».
Множественные исследования, проведенные с 1972 года, подтвердили утверждение, что «средняя доля генетических различий между людьми из разных человеческих популяций лишь немного превышает таковую между неродственными особями из одной популяции».
| Икс | Африканцы | Европейцы | Азиаты |
|---|---|---|---|
| Европейцы | 36,5 | - | - |
| Азиаты | 35,5 | 38,3 | - |
| Коренные американцы | 26,1 | 33,4 | 35 год |
Эдвардс (2003) утверждает: «Это неправда, как утверждал Nature, что« два случайных человека из одной группы почти так же отличаются, как любые два случайных человека из всего мира »», а Risch et al. (2002) заявляют: «Два кавказца более похожи друг на друга генетически, чем европеец и азиат». Однако Bamshad et al. (2004) использовали данные Rosenberg et al. (2002), чтобы исследовать степень генетических различий между людьми в континентальных группах по сравнению с генетическими различиями между людьми между континентальными группами. Они обнаружили, что, хотя этих людей можно очень точно отнести к континентальным кластерам, на индивидуальном уровне наблюдалось значительное генетическое совпадение, до такой степени, что при использовании 377 локусов отдельные европейцы примерно в 38% случаев были генетически более похожими на Восточные азиаты, чем другие европейцы.
Уизерспун и др. (2007) утверждали, что даже когда индивидов можно надежно отнести к определенным группам населения, два случайно выбранных индивида из разных популяций / кластеров могут быть более похожими друг на друга, чем на случайно выбранного члена их собственного кластера., при отборе небольшого количества SNP (как в случае с учеными Джеймсом Уотсоном, Крейгом Вентером и Сон-Джин Кимом). Они заявляют, что, используя около тысячи SNP, люди из разных популяций / кластеров никогда не становятся более похожими, что, по их словам, может показаться некоторым удивительным. Уизерспун и др. пришли к выводу, что «следует проявлять осторожность при использовании географического или генетического происхождения, чтобы делать выводы об отдельных фенотипах».
В исследовании 1994 года Кавалли-Сфорца и его коллег оценивались генетические дистанции между 42 коренными популяциями на основе 120 полиморфизмов крови. Популяции были сгруппированы в девять кластеров: африканские (к югу от Сахары), европеоидные (европейские), европеоидные (внеевропейские), северные монголоидные (за исключением арктических популяций), северо-восточные азиатские арктические, южные монголоидные (материковая и островная Юго-Восточная Азия), тихоокеанские островитянин, новогвинейцы и австралийцы, а также американцы (индейцы). Хотя кластеры демонстрируют разную степень однородности, модель с девятью кластерами представляет большинство (80 из 120) деревьев с одним признаком и полезна для демонстрации фенетических отношений между этими популяциями.
Наибольшая генетическая дистанция между двумя континентами находится между Африкой и Океанией - 0,2470. Этот показатель генетического расстояния отражает изоляцию Австралии и Новой Гвинеи с момента окончания последнего ледникового максимума, когда Океания была изолирована от материковой Азии из-за повышения уровня моря. Следующее по величине генетическое расстояние находится между Африкой и Америкой - 0,2260. Это ожидаемо, поскольку самое большое географическое расстояние по суше находится между Африкой и Южной Америкой. Наименьшее генетическое расстояние 0,0155 - между европейскими и неевропейскими европеоидами. Африка - самый генетически дивергентный континент, где все остальные группы больше связаны друг с другом, чем с африканцами к югу от Сахары. Это ожидается в соответствии с гипотезой единственного происхождения. У Европы общая генетическая изменчивость примерно в три раза меньше, чем у других континентов; генетический вклад Азии и Африки в Европу составляет соответственно две трети и одну треть.
 Кластеры генов из Розенберга (2006) для кластеров K = 7. ( Кластерный анализ делит набор данных на любое заранее заданное количество кластеров.) У людей есть гены из нескольких кластеров. Кластер, преобладающий только у калашей (желтый), отделяется только при K = 7 и выше.
Кластеры генов из Розенберга (2006) для кластеров K = 7. ( Кластерный анализ делит набор данных на любое заранее заданное количество кластеров.) У людей есть гены из нескольких кластеров. Кластер, преобладающий только у калашей (желтый), отделяется только при K = 7 и выше. Исследования генетической структуры проводятся с использованием статистических компьютерных программ, предназначенных для поиска кластеров генетически похожих людей в выборке людей. В исследованиях, подобных исследованиям Риша и Розенберга, используется компьютерная программа под названием СТРУКТУРА для поиска человеческих популяций (кластеров генов). Это статистическая программа, которая работает, помещая людей в один из произвольного числа кластеров на основе их общего генетического сходства. Многие возможные пары кластеров проверяются на каждого человека для создания нескольких кластеров. Основой для этих вычислений являются данные, описывающие большое количество однонуклеотидных полиморфизмов (SNP), генетических вставок и делеций ( инделей ), микросателлитных маркеров (или коротких тандемных повторов, STR), как они появляются у каждого из отобранных индивидов. Кластерный анализ делит набор данных на любое заранее заданное количество кластеров.
Эти кластеры основаны на нескольких генетических маркерах, которые часто являются общими для разных человеческих популяций даже в больших географических регионах. Понятие генетического кластера состоит в том, что люди в кластере в среднем разделяют одинаковые частоты аллелей друг с другом, чем у людей в других кластерах. ( AWF Edwards, 2003, но также см. Инфобокс «Multi Locus Allele Clusters»). При тестировании идеализированных популяций компьютерная программа STRUCTURE постоянно недооценивала количество популяций в наборе данных при высокой скорости миграции между популяциями и низкой скорости мутаций. (например, однонуклеотидный полиморфизм ). В 2004 году Линн Джорд и Стивен Вудинг утверждали, что «анализ многих локусов теперь дает достаточно точные оценки генетического сходства между индивидуумами, а не популяциями. Группирование индивидуумов коррелирует с географическим происхождением или родословной».
С 2002 года был проведен ряд генетических кластерных исследований, в том числе следующие:
| Авторы | Год | заглавие | Размер выборки / количество отобранных популяций | Образец | Маркеры |
| Розенберг и др. | 2002 г. | Генетическая структура популяций человека | 1056/52 | Проект разнообразия генома человека (HGDP-CEPH) | 377 STR |
| Серре и Паабо | 2004 г. | Человеческие отношения во всем мире, выведенные из общегеномных паттернов вариаций | 89/15 | а: HGDP | 20 STR |
| 90 / географически распределенные люди | b: Jorde, 1997 г. | ||||
| Розенберг и др. | 2005 г. | Клины, кластеры и влияние дизайна исследования на вывод о структуре населения | 1056/52 | Проект разнообразия генома человека (HGDP-CEPH) | 783 STR + 210 инделей |
| Ли и др. | 2008 г. | Человеческие отношения во всем мире, выведенные из общегеномных паттернов вариаций | 938/51 | Проект разнообразия генома человека (HGDP-CEPH) | 650 000 SNP |
| Тишкофф и др. | 2009 г. | Генетическая структура и история африканцев и афроамериканцев | ~ 3400/185 | HGDP-CEPH плюс 133 дополнительных африканских населения и индейцев | 1327 STR + индели |
| Xing et al. | 2010 г. | На пути к более единообразной выборке генетического разнообразия человека: исследование популяций во всем мире с помощью генотипирования с высокой плотностью | 850/40 | HapMap plus 296 человек | 250 000 SNP |
В статье 2005 года Розенберг и его команда признали, что на результаты исследования структуры человеческой популяции сильно влияет способ его разработки. Они сообщили, что количество локусов, размер выборки, географическая дисперсия выборок и предположения о корреляции частоты аллелей - все это влияет на результат исследования.
В обзоре исследований разнообразия генома человека Гвидо Барбужани и его коллеги отмечают, что различные кластерные исследования выявили разное количество кластеров с разными границами. Они пишут, что несогласованные модели генетической изменчивости и высокое внутрипопуляционное генетическое разнообразие «делают [] трудным или невозможным определить, раз и навсегда, основные генетические кластеры человечества».
Генетическая кластеризация также подверглась критике со стороны антропологов Пенсильванского университета Кеннета Вайса и Брайана Ламберта. Они утверждали, что понимание структуры человеческой популяции с точки зрения дискретных генетических кластеров искажает путь, по которому образовались различные человеческие популяции, которые расходились с общими предками в Африке. По иронии судьбы, игнорируя то, как история популяции на самом деле работает как единый процесс от общего происхождения, а не как последовательность событий творения, структурный анализ, который, кажется, представляет вариации в терминах дарвиновской эволюции, в корне не является дарвиновским ».
Главный вывод Розенберга и его коллег (2002) заключался в том, что, когда программа генерировала пять кластеров (обозначенных как K = 5), «кластеры в значительной степени соответствовали основным географическим регионам». В частности, пять кластеров соответствовали Африке, Европе, Ближнему Востоку, Центральной и Южной Азии, Восточной Азии, Океании и Северной и Южной Америке. Исследование также подтвердило предыдущий анализ, показав, что «внутрипопуляционные различия между людьми составляют от 93 до 95% генетической изменчивости; различия между основными группами составляют всего от 3 до 5%».
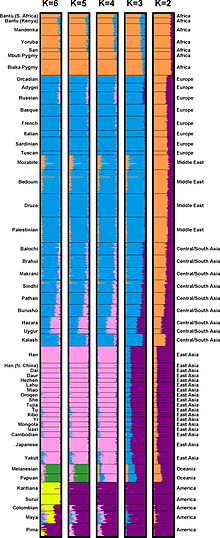 Структура популяции человека может быть выведена из данных о последовательностях мультилокусной ДНК ( Rosenberg et al. 2002, 2005). У людей из 52 популяций было исследовано 993 маркера ДНК. Эти данные были использованы для разделения людей на K = 2, 3, 4, 5 или 6 кластеров генов. На этом рисунке средняя фракционная принадлежность особей из каждой популяции представлена горизонтальными полосами, разделенными на K цветных сегментов.
Структура популяции человека может быть выведена из данных о последовательностях мультилокусной ДНК ( Rosenberg et al. 2002, 2005). У людей из 52 популяций было исследовано 993 маркера ДНК. Эти данные были использованы для разделения людей на K = 2, 3, 4, 5 или 6 кластеров генов. На этом рисунке средняя фракционная принадлежность особей из каждой популяции представлена горизонтальными полосами, разделенными на K цветных сегментов. Розенберг и его коллеги (2005) утверждали, основываясь на кластерном анализе, что популяции не всегда изменяются непрерывно, а генетическая структура популяции согласована, если включено достаточно генетических маркеров (и субъектов). "Изучение взаимосвязи между генетической и географической дистанцией поддерживает точку зрения, согласно которой кластеры возникают не как артефакт схемы выборки, а в результате небольших прерывистых скачков генетической дистанции для большинства пар популяций по разные стороны географических барьеров по сравнению с генетическими расстояние для пар на одной стороне.Таким образом, анализ набора данных с 993 локусами подтверждает наши предыдущие результаты: если достаточно маркеров используется с достаточно большой мировой выборкой, индивиды могут быть разделены на генетические кластеры, соответствующие основным географическим подразделениям земного шара, при этом некоторые люди из промежуточных географических регионов имеют смешанное членство в кластерах, соответствующих соседним регионам ". Они также писали относительно модели с пятью кластерами, соответствующими Африке, Евразии (Европа, Ближний Восток и Центральная / Южная Азия), Восточной Азии, Океании и Америке: «Для пар населения из одного кластера по мере увеличения географического расстояния генетическое расстояние увеличивается линейно в соответствии с клинальной структурой популяции. Однако для пар из разных кластеров генетическое расстояние обычно больше, чем между внутрикластерными парами, имеющими одинаковое географическое расстояние. Например, генетические расстояния для популяционных пар с одна популяция в Евразии, а другая в Восточной Азии больше, чем у пар, находящихся на эквивалентном географическом расстоянии в пределах Евразии или Восточной Азии. Грубо говоря, это эти небольшие прерывистые скачки генетической дистанции - через океаны, Гималаи и Сахару - которые обеспечивают основу для способности STRUCTURE определять кластеры, соответствующие географическим регионам ".
Розенберг заявил, что их результаты «не следует воспринимать как доказательство нашей поддержки какой-либо конкретной концепции биологической расы (...). Генетические различия между человеческими популяциями происходят в основном из-за градации частот аллелей, а не из отличительных« диагностических »генотипов». Общие результаты исследования подтвердили, что генетические различия между популяциями составляют от 93 до 95%. Между группами обнаруживается только 5% генетической изменчивости.
Исследование Розенберга подверглось критике по нескольким причинам.
Существование аллельных клин и наблюдение того, что основная часть человеческих вариаций непрерывно распределяется, привело некоторых ученых к выводу, что любая схема категоризации, пытающаяся осмысленно разделить эту вариацию, обязательно создаст искусственные усечения. (Киттлс и Вайс, 2003). Именно по этой причине, утверждает Реанна Франк, попытки распределить людей по группам предков на основе генетической информации дали разные результаты, которые сильно зависят от методологической разработки. Серр и Паабо (2004) делают аналогичное заявление:
Отсутствие сильной континентальной кластеризации в генофонде человека имеет практическое значение. Недавно было заявлено, что «самая большая генетическая структура, которая существует в человеческой популяции, происходит на расовом уровне» (Risch et al. 2002). Наши результаты показывают, что это не так, и мы не видим причин предполагать, что «расы» представляют собой какие-либо единицы, имеющие отношение к пониманию генетической истории человечества.
В ответ на Serre and Pääbo (2004) Розенберг и др. (2005) утверждают, что их кластерный анализ надежен. Кроме того, они согласны с Серром и Паабо в том, что принадлежность к нескольким кластерам может быть истолкована как свидетельство клинизма (изолированность по расстоянию), хотя они также отмечают, что это также может быть связано с примесью между соседними группами (модель малых островов). В-третьих, они отмечают, что свидетельство кластеризации не является свидетельством каких-либо конкретных концепций «биологической расы».
Кластеризация не особенно соответствует континентальному делению. В зависимости от параметров, указанных в их аналитической программе, Розенберг и Притчард смогли построить между делениями от 4 до 20 кластеров исследуемых геномов, хотя они исключили анализ с более чем 6 кластерами из своей опубликованной статьи. Значения вероятности для различных конфигураций кластера сильно различались, при этом наиболее вероятная конфигурация имела 16 кластеров, хотя другие конфигурации с 16 кластерами имели низкую вероятность. В целом, по словам генетика Деборы Болник (2008: 76-77), «нет четких доказательств того, что K = 6 было наилучшей оценкой». Количество генетических кластеров, использованных в исследовании, было выбрано произвольно. Хотя в первоначальном исследовании использовалось разное количество кластеров, в опубликованном исследовании выделено шесть генетических кластеров. Количество генетических кластеров определяется пользователем компьютерной программы, проводящей исследование. Позже Розенберг показал, что его команда использовала заранее заданное количество генетических кластеров от шести до двадцати, «но не публиковала эти результаты, потому что Structure [используемая компьютерная программа] определила несколько способов разделения выбранных особей». Дороти Робертс, профессор права, утверждает, что «в выводах группы нет ничего, что предполагало бы, что шесть кластеров представляют структуру человеческой популяции лучше, чем десять, пятнадцать или двадцать». Получив указание найти два кластера, программа определила две группы населения, привязанные к Африке и Америке. В случае шести кластеров к предыдущим пяти была добавлена вся группа калешей, этническая группа, проживающая в Северном Пакистане.
Комментируя исследование Розенберга, профессор права Дороти Робертс написала, что «исследование на самом деле показало, что существует множество способов сократить широкий диапазон генетических вариаций человека.
Сара А. Тишкофф и его коллеги проанализировали глобальную выборку, состоящую из 952 человек из исследования HGDP-CEPH, 2432 африканцев из 113 этнических групп, 98 афроамериканцев, 21 йеменца, 432 человека индийского происхождения и 10 коренных австралийцев. Глобальный анализ СТРУКТУРЫ этих людей изучил 1327 полиморфных маркеров, в том числе 848 STR, 476 инделей и 3 SNP. Авторы сообщили о кластерных результатах для K = 2 - K = 14. Внутри Африки с помощью байесовского анализа были выведены шесть групп предков, которые были тесно связаны с этнолингвистическим наследием. Популяции банту сгруппированы с другими нигеро-конго- говорящими народами из Западной Африки. Афроамериканцы в значительной степени принадлежали к этому кластеру Нигер-Конго, но также имели значительное европейское происхождение. Население Нило-Сахары сформировало свой собственный кластер. Чадское население сгруппировалось с нило-сахарскими группами, что позволяет предположить, что большинство современных носителей чадского языка первоначально говорили на языках нило-сахарской семьи, а затем приняли афро-азиатские языки. Нилотские популяции из Великих африканских озер в значительной степени принадлежали и к этому нило-сахарскому кластеру, но также имели некоторое афро-азиатское влияние из-за ассимиляции кушитских групп за последние 3000 лет. Популяции койсанов сформировали свой собственный кластер, который ближе всего сгруппировался с кластером пигмеев. На « Цветном мысе» были показаны задания из койсанского, европейского и других кластеров из-за смешанного наследия населения. В хадзе и сандаве население создало свой собственный кластер. Также был выявлен афро-азиатский кластер с афро-азиатскими носителями из Северной Африки и Африканского Рога, образующими непрерывную группу. Афро-азиатские носители в районе Великих озер в значительной степени принадлежали к этому афро-азиатскому кластеру, но также имели некоторое влияние банту и нилотов из-за ассимиляции соседних групп за последние 3000 лет. Остальные предполагаемые группы предков были связаны с европейским, ближневосточным, океаническим, индийским, индейским и восточноазиатским населением.
Цзиньчуань Син и его коллеги использовали альтернативный набор данных генотипов человека, включая образцы HapMap и их собственные образцы (296 новых особей из 13 популяций), в общей сложности 40 популяций, распределенных примерно равномерно по поверхности земли. Они обнаружили, что альтернативная выборка снизила оценку F ST межпопуляционных различий с 0,18 до 0,11, предполагая, что более высокое число может быть артефактом неравномерной выборки. Они провели кластерный анализ с использованием программы ADMIXTURE и обнаружили, что «генетическое разнообразие распределяется по более клинической схеме, когда выбираются более географически промежуточные популяции».
Исследование, проведенное Паназиатским консорциумом SNP HUGO в 2009 году с использованием аналогичного анализа основных компонентов, показало, что популяции Восточной и Юго-Восточной Азии сгруппированы вместе, и предположило общее происхождение этих популяций. В то же время они наблюдали значительный разрыв между этим кластером и Южной Азией, комментируя, что «большинство индейских популяций демонстрировали свидетельства общего происхождения с европейскими популяциями». Было отмечено, что «генетическая родословная сильно коррелирует с лингвистической принадлежностью, а также географией».
Исследования кластеризации вновь открыли дискуссию о научной реальности расы или ее отсутствии. В конце 1990-х годов эволюционный генетик из Гарварда Ричард Левонтин заявил, что «нельзя предложить никаких оправданий для продолжения биологической концепции расы. (...) Генетические данные показывают, что независимо от того, как расовые группы определяются, два человека из одной расовой группы являются примерно так же отличаются друг от друга, как два человека из любых двух разных расовых групп. С тех пор эта точка зрения была подтверждена многочисленными авторами и Американской ассоциацией физических антропологов. AWF Эдвардс, а также Рик Киттлс и Джеффри Лонг критиковали методологию Левонтина вместе с Лонгом. отмечая, что между людьми и шимпанзе больше сходства, чем различий, и больше генетических вариаций у шимпанзе и людей, чем между ними. Эдвардс также заявил, что Левонтин совершил «неоправданное нападение на человеческую классификацию, о чем он сожалел по социальным причинам». статьи, Кейт Ханли, Грасиела Кабана и Джеффри Лонг пересчитывают распределение человеческого разнообразия, используя более сложную lex, чем Левонтин и его последователи. Они приходят к выводу: «В общем, мы согласны с выводом Левонтина о том, что основанные на западе расовые классификации не имеют таксономического значения, и мы надеемся, что это исследование, которое принимает во внимание наше текущее понимание структуры человеческого разнообразия, дает его основополагающее открытие более надежным. эволюционная основа ".
Исследования генетической кластеризации, и особенно результат пяти кластеров, опубликованный командой Розенберга в 2002 году, интерпретировались журналистом Николасом Уэйдом, эволюционным биологом Арманом Мари Леруа и другими как демонстрация биологической реальности расы. Для Леруа «Раса - это просто сокращение, которое позволяет нам разумно, хотя и без большой точности, говорить о генетических, а не культурных или политических различиях». Он заявляет, что «можно разделить население мира на 10, 100, возможно, 1000 групп», и описывает европейцев, басков, жителей Андаманских островов, ибосов и кастильцев как «расу». В ответ на заявления Леруа Совет по исследованиям в области социальных наук созвал группу экспертов для обсуждения расы и геномики в Интернете. В своих статьях 2002 и 2005 годов Розенберг и его коллеги не согласны с тем, что их данные подразумевают биологическую реальность расы.
В 2006 году Левонтин написал, что любое генетическое исследование требует некоторой априорной концепции расы или этнической принадлежности, чтобы упаковать генетическое разнообразие человека в определенное, ограниченное количество биологических групп. Получив информацию от генетики, зоологи давно отказались от концепции расы для разделения групп не относящихся к человеку популяций животных внутри вида. При определении по разным критериям у одного и того же вида можно было выделить большое количество рас. Левонтин отмечает, что генетическое тестирование показало, что «поскольку оказалось, что многие из этих рас основаны только на одном или двух генах, два животных, рожденные в одном помете, могут принадлежать к разным« расам »».
Исследования, направленные на поиск генетических кластеров, информативны настолько, насколько информативны популяции, которые они отбирают. Например, Риш и Бурхард полагались на две или три местных популяции с пяти континентов, которые вместе должны были представлять весь человеческий род. В другом исследовании генетической кластеризации для представления Африки использовались три группы населения к югу от Сахары; Образцы из Китая, Японии и Камбоджи для Восточной Азии; Североевропейские и северные итальянские образцы для обозначения «кавказцев». Целые регионы, субконтиненты и суши не участвуют во многих исследованиях. Кроме того, не были определены социально-географические категории, такие как «Восточная Азия» и «кавказцы». «Горстка этнических групп, символизирующих целый континент, подражает основному принципу расового мышления: поскольку расы состоят из однородных людей, любой может представлять всю группу», - отмечает Робертс.
Модель Big Few не работает, если включить в нее упущенные из виду географические регионы, такие как Индия. Исследование 2003 года, в ходе которого были изучены пятьдесят восемь генетических маркеров, показало, что индейские народы обязаны своим наследственным происхождением Африке, Центральной Азии, Европе и южному Китаю. Рирдон из Принстонского университета утверждает, что ошибочные методы отбора проб являются неотъемлемой частью многих проектов генетических исследований. Проект разнообразия генома человека (HGDP) основывался на образцах, которые, как предполагалось, были географически разделены и изолированы. Относительно небольшие размеры выборки коренных популяций для HGDP не отражают генетического разнообразия человеческого вида и не отражают миграции и смешение популяционных групп, которые происходили с доисторических времен. Географические области, такие как Балканы, Ближний Восток, Северная и Восточная Африка и Испания, редко включаются в генетические исследования. Например, коренное население Восточной и Северной Африки никогда не отбирается для представления Африки, потому что оно не соответствует профилю «черной» Африки. Предполагается, что отобранные коренные популяции HGDP являются «чистыми»; профессор права Робертс утверждает, что «их необычайная чистота - это еще одна причина, по которой они не могут заменить все другие народы мира, отмеченные смешением в результате миграции, торговли и завоеваний».
Кинг и Мотульски в статье Science 2002 года заявляют, что «Хотя компьютерные данные всех этих исследований предлагают более глубокое понимание генетического единства и разнообразия человеческого вида, а также его древней миграционной истории, ни одно из них не поддерживает разделение виды на дискретные, генетически детерминированные расовые категории ". Кавалли-Сфорца утверждает, что отнесение кластеров к расам было бы «бесполезным занятием», потому что «каждый уровень кластеризации будет определять другую популяцию, и нет биологической причины отдавать предпочтение определенной группе». Бамшад в статье 2004 года, опубликованной в журнале Nature, утверждает, что для более точного изучения генетической изменчивости человека будет использоваться метод объективной выборки, который будет выбирать популяции случайным образом и систематически по всему миру, включая те популяции, которые характеризуются историческим смешением, а не вишневым. -выбор выборок населения, которые соответствуют априорным представлениям о расовой классификации. Робертс заявляет, что «если бы исследователи собирали образцы ДНК непрерывно от региона к региону по всему миру, они сочли бы невозможным установить четкие границы между большими географическими группами».
Антропологи, такие как К. Лоринг Брейс, философы Джонатан Каплан и Расмус Винтер и генетик Джозеф Грейвс, утверждали, что, хотя, безусловно, можно найти биологические и генетические вариации, которые примерно соответствуют группировкам, обычно определяемым как «континентальные расы», это верно почти для всех географически различных групп населения. Таким образом, кластерная структура генетических данных зависит от исходных гипотез исследователя и отобранных популяций. При отборе образцов континентальных групп группы становятся континентальными; если бы кто-то выбрал другие образцы выборки, кластеризация была бы другой. Вайс и Фуллертон отметили, что если взять образцы только исландцев, майя и маори, то сформируются три отдельных кластера, и все остальные популяции можно будет описать как клинически состоящие из примесей генетических материалов маори, исландцев и майя. Таким образом, Каплан и Винтер утверждают, что Левонтин и Эдвардс правы в своих аргументах. Они пришли к выводу, что, хотя расовые группы характеризуются разными частотами аллелей, это не означает, что расовая классификация является естественной таксономией человеческого вида, потому что в человеческих популяциях можно найти множество других генетических паттернов, которые пересекают расовые различия. Более того, геномные данные недооценивают, хотят ли мы видеть подразделения (т. Е. Сплиттеры) или континуум (т. Е. Комочники). Согласно точке зрения Каплана и Винтера, расовые группировки являются объективными социальными конструкциями (см. Mills 1998), которые имеют условную биологическую реальность только постольку, поскольку категории выбираются и конструируются по прагматическим научным причинам.
Коммерческие компании по тестированию родословных, использующие данные генетической кластеризации, также подверглись резкой критике. Ограничения генетической кластеризации усиливаются, когда предполагаемая популяционная структура применяется к индивидуальному происхождению. Тип статистического анализа, проводимого учеными, плохо влияет на индивидуальное происхождение, потому что они смотрят на разницу в частотах, а не на абсолютные различия между группами. Коммерческие компании, занимающиеся генетической генеалогией, виновны в том, что Пиллар Оссорио называет «тенденцией превращать статистические утверждения в категоричные». Не только люди одной и той же местной этнической группы, но и два брата и сестры могут оказаться членами разных континентальных групп или «рас» в зависимости от наследуемых аллелей.
Многие коммерческие компании используют данные из первоначальной фразы Международного проекта HapMap (HapMap), в которой образцы населения были собраны из четырех этнических групп в мире: китайцев хань, японцев, йоруба-нигерийцев и жителей Юты североевропейского происхождения. Если у человека есть предки из региона, где в компьютерной программе нет образцов, он будет компенсировать ближайшим образцом, который может не иметь ничего общего с фактическим происхождением клиента: «Рассмотрим тестирование генетического происхождения, проведенное на человеке, которого мы назовем Джо, восемь прабабушек и дедушек которого были из южной Европы. Популяции HapMap используются в качестве эталонов для проверки генетической родословной Джо. Европейские образцы HapMap состоят из "северных" европейцев. В регионах генома Джо, которые различаются между северной и южной Европой (такие регионы может включать ген лактазы), тест на генетическое происхождение, использующий эталонную популяцию HapMap, вероятно, неправильно отнесет происхождение этой части генома к неевропейской популяции, потому что эта геномная область будет больше похожа на Yoruba HapMap или ханьских китайских образцов, чем образцы из Северной Европы. Точно так же человек, имеющий западноевропейское и западноафриканское происхождение, может иметь Наследники из Западной Европы и Западной Африки, или вместо этого будут назначены в Восточную Африку, где могут быть найдены различные предки. "Сообщение покупателям о том, что они являются составной частью нескольких антропологических группировок, усиливает три основных мифа о расе: что существуют чистые расы, что каждая раса содержит людей, которые в своей основе идентичны и фундаментально отличаются от людей других рас, и что расы могут быть биологически идентичными. разграничены ". Многие компании основывают свои выводы на неадекватных и ненаучных методах отбора проб. Исследователи никогда не отбирали выборки среди населения мира систематическим и случайным образом.
Робертс выступает против использования широких географических или континентальных группировок: «молекулярные генетики обычно относятся к африканскому происхождению так, как будто все на континенте больше похожи друг на друга, чем на людей других континентов, которые могут быть ближе как географически, так и генетически. Эфиопы имеют более близкое генетическое родство с армянами, чем с популяциями банту. Точно так же сомалийцы генетически более схожи с популяциями арабских стран Персидского залива, чем с другими популяциями в Африке. Браун и Хаммондс (2008) утверждают, что неправильное восприятие континентов как естественных групп населения коренится в предположение, что популяции являются естественными, изолированными и статичными. Популяции стали рассматриваться как «ограниченные единицы, поддающиеся научному отбору, анализу и классификации». Человеческие существа по своей природе не организованы в определяемые генетически сплоченные популяции.
Программное обеспечение, поддерживающее расчет генетической кластеризации.