
Войти
| Гомосерин дегидрогеназа | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Комплекс гомосериндегидрогеназы с аналогом НАД + и L-гомосерином. Комплекс гомосериндегидрогеназы с аналогом НАД + и L-гомосерином. | |||||||||
| Идентификаторы | |||||||||
| Условное обозначение | Homoserine_dh | ||||||||
| Pfam | PF00742 | ||||||||
| ИнтерПро | IPR001342 | ||||||||
| ПРОФИЛЬ | PDOC00800 | ||||||||
| SCOP2 | 1ebu / SCOPe / SUPFAM | ||||||||
| |||||||||
| Гомосерин дегидрогеназа | |||||||||
|---|---|---|---|---|---|---|---|---|---|
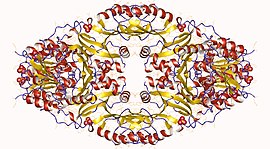 Гомотетрамер гомосериндегидрогеназы, Thiobacillus denitrificans Гомотетрамер гомосериндегидрогеназы, Thiobacillus denitrificans | |||||||||
| Идентификаторы | |||||||||
| ЕС нет. | 1.1.1.3 | ||||||||
| № CAS | 9028-13-1 | ||||||||
| Базы данных | |||||||||
| IntEnz | Просмотр IntEnz | ||||||||
| BRENDA | BRENDA запись | ||||||||
| ExPASy | Просмотр NiceZyme | ||||||||
| КЕГГ | Запись в KEGG | ||||||||
| MetaCyc | метаболический путь | ||||||||
| ПРИАМ | профиль | ||||||||
| Структуры PDB | RCSB PDB PDBe PDBsum | ||||||||
| Генная онтология | Amigo / QuickGO | ||||||||
| |||||||||
В энзимологии, A гомосериндегидрогеназа ( ЕС 1.1.1.3 ) представляет собой фермент, который катализирует в химическую реакцию
2 субстрата этого фермента - это L-гомосерин и NAD + (или NADP + ), тогда как его 3 продукта - это L-аспартат, 4-полуальдегид, NADH (или NADPH ) и H +.
Этот фермент принадлежит к семейству оксидоредуктаз, особенно тех, которые действуют на группу CH-OH донора с НАД + или НАДФ + в качестве акцептора. Систематическое название данного фермента класс L-гомосерин: NAD (P) + оксидоредуктаза. Другие широко используемые имена включают HSDH и HSD.
Гомосериндегидрогеназа катализирует третью стадию аспартатного пути; NAD (P) -зависимое снижение аспартат - беты-полуальдегид в гомосерина. Гомосерин является промежуточным продуктом в биосинтезе из треонина, изолейцина и метионина.
Фермент можно найти в монофункциональной форме у некоторых бактерий и дрожжей. Структурный анализ монофункционального фермента дрожжей показывает, что фермент представляет собой димер, состоящий из трех отдельных областей; N-концевой нуклеотид-связывающий домен, короткая центральная димеризационная область и C-концевой каталитический домен. N-концевой домен образует модифицированную складку Россмана, в то время как каталитический домен образует новый смешанный альфа-бета-лист.
Фермент также может быть найден в бифункциональной форме, состоящей из N-концевого домена аспартокиназы и C-концевого домена гомосериндегидрогеназы, как у бактерий, таких как Escherichia coli, и у растений.
Бифункциональный фермент аспартокиназа-гомосериндегидрогеназа (AK-HSD) имеет регуляторный домен, который состоит из двух субдоменов с общей петлей - альфа-спираль- петля- бета-цепь петля-бета-цепь. Каждый субдомен содержит домен ACT, который позволяет комплексно регулировать несколько различных функций белка. Ген AK-HSD кодирует аспартаткиназу, промежуточный домен (кодирующий линкерную область между двумя ферментами в бифункциональной форме) и, наконец, последовательность, кодирующую гомосериндегидрогеназу.
По состоянию на конец 2007 г. для этого класса ферментов были решены 4 структуры с кодами доступа 1EBF, 1EBU, 1Q7G и 1TVE.
 Рис. 2. Мультяшное изображение активного сайта гомосериндегидрогеназы ( PBD 1EBU ).
Рис. 2. Мультяшное изображение активного сайта гомосериндегидрогеназы ( PBD 1EBU ). Гомосериндегидрогеназа катализирует реакцию полуальдегида аспартата (ASA) с гомосерином. Общая реакция восстанавливает функциональную группу C4 карбоновой кислоты ASA до первичного спирта и окисляет альдегид C1 до карбоновой кислоты. Считается, что остатки Glu 208 и Lys 117 вовлечены в активный каталитический сайт фермента. Было показано, что Asp 214 и Lys 223 важны для переноса гидрида в катализируемой реакции.
После того, как С4 - карбоновая кислота является уменьшена до альдегида, и C1 - альдегид окисляется до карбоновой кислоты, эксперименты показывают, что Asp 219, Glu 208 и молекула воды связывает ASA в активном центре, а Lys 223 жертвует протон к аспартатам-полуальдегиду Кислород C4. Гомосериндегидрогеназа имеет кофактор NAD (P) H, который затем отдает водород тому же углероду, эффективно восстанавливая альдегид до спирта. (См. Рисунки 1 и 2).
Однако точный механизм полного катализа гомосериндегидрогеназы остается неизвестным.
Постулируется, что реакция, катализируемая гомосериндегидрогеназой, протекает по би-би- кинетическому механизму, при котором кофактор НАД (Ф) Н первым связывает фермент и последним отделяется от фермента после завершения реакции. Кроме того, хотя и НАДН, и НАДФН являются подходящими кофакторами для реакции, НАДН является предпочтительным. К м реакции в четыре раза меньше с NADH и К кошки / К м в три раза больше, что указывает на более эффективную реакцию.
Гомосериндегидрогеназа также демонстрирует кинетику разного порядка при субстратных уровнях субстрата. Кроме того, вариабельная кинетика гомосериндегидрогеназы является артефактом более быстрой диссоциации аминокислотного субстрата из ферментного комплекса по сравнению с диссоциацией кофактора.
Путь метаболизма аспартата участвует как в хранении аспарагина, так и в синтезе аминокислот семейства аспартатов. Гомосериндегидрогеназа катализирует промежуточную стадию в этом пути хранения и использования азота и углерода. (См. Рисунок 3).
В фотосинтезирующих организмах глутамин, глутамат и аспартат накапливаются в течение дня и используются для синтеза других аминокислот. Ночью аспартат превращается в аспарагин для хранения. Кроме того, ген аспартаткиназы-гомосериндегидрогеназы в первую очередь экспрессируется в активно растущих молодых тканях растений, особенно в апикальной и латеральной меристемах.
У млекопитающих отсутствуют ферменты, участвующие в метаболическом пути аспартата, включая гомосериндегидрогеназу. Поскольку лизин, треонин, метионин и изолейцин вырабатываются этим путем, они считаются незаменимыми аминокислотами для млекопитающих.
 Рисунок 3. Гомосериндегидрогеназа - это фермент, участвующий в пути биосинтеза нескольких ключевых аминокислот. Он негативно регулируется треонином, и этот путь подлежит дополнительной регуляции.
Рисунок 3. Гомосериндегидрогеназа - это фермент, участвующий в пути биосинтеза нескольких ключевых аминокислот. Он негативно регулируется треонином, и этот путь подлежит дополнительной регуляции. Гомосериндегидрогеназа и аспартаткиназа подлежат значительному регулированию (см. Рисунок 3). HSD ингибируется продуктами метаболического пути аспартата, в основном треонином. Треонин действует как конкурентный ингибитор как HSD, так и аспартаткиназы. У организмов, экспрессирующих AK-HSD, один из сайтов связывания треонина находится в линкерной области между AK и HSD, что свидетельствует о потенциальном аллостерическом ингибировании обоих ферментов.
Однако существуют некоторые устойчивые к треонину формы HSD, для ингибирования которых требуются концентрации треонина, намного превышающие физиологически присутствующие. Эти нечувствительные к треонину формы HSD используются в генно-инженерных растениях для увеличения выработки треонина и метионина для повышения питательной ценности.
Гомосериндегидрогеназа также регулируется транскрипцией. Его промоторная последовательность содержит последовательность цис-регуляторного элемента TGACTC, которая, как известно, участвует в других путях биосинтеза аминокислот. Opaque2 регуляторный элемент также участвует в регуляции гомосериндегидрогеназу дегидрогеназы, но его последствия до сих пор не определены.
У растений также существует регуляция экспрессии гена AK-HSD в окружающей среде. Было продемонстрировано, что воздействие света увеличивает экспрессию гена AK-HSD, предположительно связанного с фотосинтезом.
У людей наблюдается значительный рост заболеваемости патогенными грибами, поэтому разработка противогрибковых препаратов является важной биохимической задачей. Поскольку гомосериндегидрогеназа обнаруживается в основном в растениях, бактериях и дрожжах, но не в млекопитающих, она является серьезной мишенью для разработки противогрибковых препаратов. Недавно было обнаружено, что 5-гидрокси-4-оксонорвалин (HON) необратимо направляет и подавляет активность HSD. HON структурно подобен полуальдегиду аспартата, поэтому предполагается, что он служит конкурентным ингибитором HSD. Аналогичным образом, (S) 2-амино-4-оксо-5-гидроксипентановая кислота (RI-331), другой аналог аминокислоты, также, как было показано, ингибирует HSD. Оба эти соединения эффективны, среди прочего, против Cryptococcus neoformans и Cladosporium fulvum.
В дополнение к аналогам аминокислот, несколько фенольных соединений, как было показано, ингибируют активность HSD. Подобно HON и RI-331, эти молекулы являются конкурентными ингибиторами, которые связываются с активным центром фермента. В частности, фенольная гидроксильная группа взаимодействует с сайтом связывания аминокислоты.
