
Войти
 Гиппокамп человека.
Гиппокамп человека.  Окрашенный по Нисслю коронарный срез мозга макаки с гиппокампом (обведен). Источник: brainmaps.org
Окрашенный по Нисслю коронарный срез мозга макаки с гиппокампом (обведен). Источник: brainmaps.org Анатомия гиппокампа описывает физические аспекты и свойства гиппокампа, нервной структуры в медиальной височной доле мозга. Он имеет характерную изогнутую форму, которую сравнивают с морским коньком чудовищем греческой мифологии и рогами барана Амона в египетской мифологии.. Эта общая схема распространяется на весь спектр видов млекопитающих, от ежа до человека, хотя детали меняются. Например, у крысы два гиппокампа похожи на пару бананов, соединенных стеблями. В мозге приматов, включая людей, часть гиппокампа у основания височной доли намного шире, чем часть вверху. Из-за трехмерной кривизны этой конструкции обычно видны двумерные сечения, подобные показанным. Нейровизуализация изображения могут отображать различные формы в зависимости от угла и местоположения разреза.
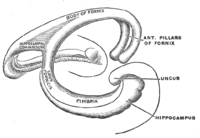 Форма человеческого гиппокампа и связанных с ним структур.
Форма человеческого гиппокампа и связанных с ним структур. Топологически поверхность полушария головного мозга можно рассматривать как сферу с углублением в месте прикрепления к среднему мозгу. Структуры, выстилающие край отверстия, вместе составляют так называемую лимбическую систему (латинское limbus = граница), причем гиппокамп выстилает задний край этого отверстия. Эти лимбические структуры включают гиппокамп, поясную извилину, обонятельную кору и миндалину. Пол Маклин однажды предположил в рамках своей теории триединого мозга, что лимбические структуры составляют нейронную основу эмоции. Хотя большинство нейробиологов больше не верят в концепцию единой «лимбической системы», эти области сильно взаимосвязаны и взаимодействуют друг с другом.
 Основная схема гиппокампа, показанная с использованием модифицированного рисунка Рамона-и-Кахала. ДГ: зубчатая извилина. Sub: subiculum. ЭК: энторинальная кора
Основная схема гиппокампа, показанная с использованием модифицированного рисунка Рамона-и-Кахала. ДГ: зубчатая извилина. Sub: subiculum. ЭК: энторинальная кора Начало от зубчатой извилины и движение внутрь вдоль S-образной кривой гиппокампа означает прохождение ряда узких зон. Первая из них, зубчатая извилина (DG), на самом деле представляет собой отдельную структуру, плотно упакованный слой мелких гранулярных клеток, обернутых вокруг конца собственно гиппокампа, образуя в одних сечениях заостренный клин, в других - полукруг. Затем идет ряд областей Cornu Ammonis: сначала CA4 (который лежит в основе зубчатой извилины), затем CA3, затем очень маленькая зона под названием CA2, затем CA1. Все области CA заполнены плотно упакованными пирамидными клетками, подобными тем, которые обнаруживаются в неокортексе. После CA1 идет область, называемая subiculum. После этого идет пара нечетко очерченных областей, называемых предубикулумом и парапубикулумом, а затем переход к собственно коре (в основном, энторинальной области коры). Большинство анатомов используют термин «собственно гиппокамп» для обозначения четырех полей СА, и образование гиппокампа для обозначения собственно гиппокампа плюс зубчатая извилина и субикулум.
Основная передача сигналов пути проходят через гиппокамп и объединяются, образуя петлю. Большая часть внешнего входа поступает из прилегающей энторинальной коры через аксоны так называемого перфорантного пути. Эти аксоны возникают из слоя 2 энторинальной коры (ЭК) и заканчиваются зубчатой извилиной и СА3. Существует также отдельный путь от слоя 3 EC непосредственно к CA1, часто называемый темпорааммоническим путем или путем TA-CA1. Гранулярные клетки DG отправляют свои аксоны (так называемые «мшистые волокна») к CA3. Пирамидные клетки CA3 отправляют свои аксоны в CA1. Пирамидные клетки CA1 отправляют свои аксоны в субикулюм и глубокие слои ЭК. Субикулярные нейроны посылают свои аксоны в основном в ЭК. Перфорантный путь от зубчатой извилины к CA3-к-CA1 был назван трисинаптической цепью Пером Андерсеном, который отметил, что тонкие срезы можно вырезать из гиппокампа перпендикулярно его длинной оси, в способ, который сохраняет все эти связи. Это наблюдение легло в основу его гипотезы о ламеллярности, согласно которой гиппокамп можно рассматривать как серию параллельных полос, действующих функционально независимым образом. Пластинчатая концепция до сих пор иногда считается полезным организующим принципом, но более свежие данные, показывающие обширные продольные связи внутри системы гиппокампа, потребовали ее существенной модификации.
Вход перфорантного пути из слоя EC II входит. зубчатая извилина и передается в область СА3 (и к мшистым клеткам, расположенным в воротах зубчатой извилины, которые затем отправляют информацию в отдаленные части зубчатой извилины, где цикл повторяется). Область CA3 объединяет этот вход с сигналами от уровня EC II и отправляет обширные соединения в пределах области, а также отправляет соединения к strata radiatum и oriens ипсилатеральной и контралатеральной областей CA1 через набор волокон, называемых коллатералями Шаффера, и комиссуральный путь соответственно. Область CA1 получает входные данные из подполя CA3, слоя III EC и nucleus reuniens таламуса (которые проецируются только на концевые апикальные дендритные пучки в stratum lacunosum-molculare ). В свою очередь, CA1 проецирует на субикулум, а также отправляет информацию по вышеупомянутым выходным путям гиппокампа. Субикулум - это последняя стадия пути, объединяющая информацию от проекции CA1 и слоя III EC, чтобы также отправлять информацию по выходным путям гиппокампа.
Гиппокамп также получает ряд подкорковых входов. В Macaca fascicularis эти входные данные включают миндалевидное тело (в частности, переднюю миндалевидную область, базолатеральное ядро и периамигдалоидную кору), медиальную перегородку и диагональная полоса Брока, клауструм, субстанция innominata и базальное ядро Мейнерта, таламус ( включая передний ядерный комплекс, латеродорсальное ядро, паравентрикулярное и паратениальное ядра, ядро reuniens и центральное ядро медиалис), латеральную преоптическую и латеральную гипоталамическую области, супрамаммиллярную и ретромаммиллярную области, вентральная тегментальная область, тегментальные ретикулярные поля, ядра шва (центральное ядро шва и дорсальное ядро шва), nucleus reticularis tegementi pontis, периакведуктальный серый, дорсальное тегментальное ядро и голубое пятно. Гиппокамп также получает прямые моносинаптические проекции от мозжечка фастигиального ядра.
Эти волокна начинаются с вентральной части энторинальной коры (EC) и содержат комиссуральные (EC ◀ ▶ Hippocampus) и перфорантные (возбуждающие EC ▶ CA1 и тормозные EC ▶ CA2) волокна. Они перемещаются вдоль перегородочно-височной оси гиппокампа. Волокна перфорантного пути, как следует из названия, перфорируют субикулюм перед тем, как попасть в гиппокамп (поля СА) и зубчатую извилину.
 Венечный разрез нижнего рога бокового желудочка. (Фимбрия помечена слева в центре, а альвеус справа).
Венечный разрез нижнего рога бокового желудочка. (Фимбрия помечена слева в центре, а альвеус справа). Волокна фимбрии и свода - это гиппокамп и субикулярные ворота в и из подкорковых областей мозга. Разным частям этой системы даны разные названия:
На уровне контура альвеус содержит аксонные волокна от DG и от пирамидных нейронов CA3, CA2, CA1 и субикулума (CA1 ▶ subiculum и CA1 ▶ энторинальный проекции), которые накапливаются в височном гиппокампе, образуя фимбрию / свод, один из основных выходов гиппокампа. У крысы некоторые медиальные и латеральные энторинальные аксоны (энторинальная ▶ CA1 проекция) проходят через альвеус по направлению к молекуле CA1 stratum lacunosum, не образуя значительного количества проходящих бутонов на других слоях CA1 (Temporoammonic alvear путь ). Контралатеральные энторинальные ▶ выступы CA1 почти исключительно проходят через альвеус. Чем больше перегородка, тем больше ипсилатеральных проекций энторинального CA1, которые проходят альвеарным путем (вместо перфорантного пути). Хотя субикулум посылает аксональные проекции в альвеус, субикулум ▶ Проекция CA1 проходит через ориентировочные слои и молекулы субикулума и CA1. Холинергические и ГАМКергические проекции от MS-DBB к CA1 также проходят через Fimbria. Стимуляция фимбрии приводит к холинергическому возбуждению клеток CA1 O-LMR.
Также известно, что внеклеточная стимуляция фимбрии стимулирует пирамидные клетки CA3 антидромно и ортодромно, но не влияет на клетки зубчатых гранул. Каждая пирамидная клетка CA1 также отправляет аксональную ветвь к фимбриям.
Мшистые клетки корня и пирамидные клетки CA3 являются основными источниками комиссуральных волокон гиппокампа . Они проходят через гиппокампальные спайки, достигая контралатеральных областей гиппокампа. Комиссуры гиппокампа имеют дорсальный и вентральный сегменты. Дорсальные комиссуральные волокна состоят в основном из энторинальных и предубикулярных волокон, ведущих к или от гиппокампа и зубчатой извилины. На практике можно сказать, что каждое цитоархитектоническое поле, которое вносит вклад в комиссуральную проекцию, также имеет параллельное ассоциативное волокно, которое заканчивается в ипсилатеральном гиппокампе. Внутренний молекулярный слой зубчатой извилины (дендриты как гранулярных клеток, так и ГАМКергических интернейронов) получает проекцию, которая имеет как ассоциативные, так и комиссуральные волокна в основном из прикорневых мшистых клеток и в некоторой степени из пирамидных клеток CA3c. Поскольку эти проекционные волокна берут начало как с ипсилатеральной, так и с контралатеральной сторон гиппокампа, они называются ассоциативными / комиссуральными проекциями. Фактически, каждая мшистая клетка иннервирует как ипсилатеральную, так и контрлатеральную зубчатую извилину. Хорошо известный трисинаптический контур гиппокампа проходит в основном горизонтально вдоль гиппокампа. Однако ассоциативные / комиссуральные волокна, такие как ассоциативные проекции пирамидных клеток CA2, простираются в основном продольно (дорсовентрально) вдоль гиппокампа. Комиссуральные волокна, которые происходят из пирамидных клеток CA3, идут в области CA3, CA2 и CA1. Подобно мшистым клеткам, одна пирамидная клетка CA3 вносит вклад как в комиссуральные, так и в ассоциативные волокна, и они оканчиваются как на основных клетках, так и на интернейронах. Слабый комиссуральный выступ соединяет вместе обе области CA1. Субикулум не имеет комиссуральных входов и выходов. По сравнению с грызунами комиссуральные связи гиппокампа у обезьян и людей гораздо менее распространены. Хотя возбуждающие клетки вносят основной вклад в комиссуральные пути, сообщалось о ГАМКергическом компоненте среди их окончаний, происхождение которых восходит к воротам. Стимуляция комиссуральных волокон антидромно стимулирует DG внутригрудные перфорантные пути (HIPP) и CA3 триламинарные клетки.
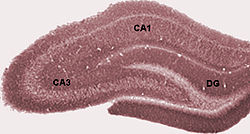 Фотография областей гиппокампа в крысиный мозг. DG: Зубчатая извилина.
Фотография областей гиппокампа в крысиный мозг. DG: Зубчатая извилина. Схема, показывающая области собственно гиппокампа по отношению к другим структурам.
Схема, показывающая области собственно гиппокампа по отношению к другим структурам. Собственно гиппокамп состоит из ряда подполей. Хотя терминология у разных авторов различается, наиболее часто используются термины зубчатая извилина и рога аммониса (буквально «рога Амона », сокращенно CA ). Зубчатая извилина содержит зубчатую фасцию и ворот, в то время как СА разделена на поля СА1, СА2, СА3 и СА4.
. Однако область, известная как CA4 на самом деле является «глубоким полиморфным слоем зубчатой извилины» (как пояснил Теодор Блэкстад (1956) и Дэвид Амарал (1978)).
Разрез поперечный разрез, гиппокамп - это С-образная структура, напоминающая рога рама . Название Cornu Ammonis относится к египетскому божеству Амону, у которого голова барана. Рогатый вид гиппокампа обусловлен различиями в плотности клеток и наличием в различной степени нейрональных волокон.
У грызунов гиппокамп расположен примерно так, что один конец находится около вершины головы (спинной или перегородочный конец), а другой конец - около низа головы (брюшной или височный конец).. Как показано на рисунке, сама структура изогнута, а подполя или области определены вдоль кривой от CA4 до CA1 (помечены только CA3 и CA1). Области CA также структурированы по глубине в четко определенные слои (или слои):
зубчатая извилина состоит из аналогичного ряда слоев:
Для современной «базы знаний» о типах нейронов гиппокампа, их профилях биомаркеров, активных и пассивных электрофизиологических параметрах, а также возможности подключения можно найти на Hippocampome.org.