принцип в генетике

Пропорции Харди - Вайнберга для двух
аллелей : горизонтальная ось показывает две
частоты аллелей p и q, а вертикальная ось показывает ожидаемые частоты
генотипов. Каждая линия показывает один из трех отдельных генотипов.
В популяционной генетике используется принцип Харди - Вайнберга, также известный как равновесие Харди - Вайнберга, модель, теорема, или закон, гласит, что частоты аллелей и генотипов в процессе роста остаются постоянными от поколения к поколению в отсутствие других эволюционных влияний. Эти влияния включают генетический дрейф, выбор партнера, ассортативное спаривание, естественный отбор, половой отбор, мутация, поток генов, мейотический драйв, генетический автостоп, узкое место популяции, эффект основателя и инбридинг.
В простейшем случае одного локуса с двумя аллелями, обозначенными A и a, с частотами f(A) = p и f(a) = q, соответственно ожидаемые частоты генотипов при случайном скрещивании равны f(AA) = p для AA гомозигот, f(aa) = q для гомозигот aa, и f(Aa) = 2pq для гетерозигот. В отсутствие отбора, мутации, генетического дрейфа или других частотах аллелей p и q постоянны между поколениями, поэтому равновесие достигается.
Принцип назван в честь G. Х. Харди и Вильгельм Вайнберг, которые осуществляли это математически. Статья Харди была сосредоточена на опровержении широко распространенного тогда мнения о том, что доминантный аллель автоматически имеет тенденцию к увеличению частоты; сегодня путаница между доминированием и отбором встречается реже. Сегодня тесты на частоту генотипов Харди - Вайнберга используются в основном для проверки стратификации населения и других форм неслучайного спаривания.
Содержание
- 1 Деривация
- 2 Отклонения от равновесия Харди - Вайнберга
- 3 Половая связь
- 4 Обобщения
- 4.1 Обобщение для более чем двух аллелей
- 4.2 Обобщение для полиплоидии
- 4.3 Полное обобщение
- 5 Тесты значимости отклонения
- 5.1 Пример
 тест отклонения
тест отклонения - 5.2 Точный тест Фишера (вероятностный тест)
- 6 Тесты на эквивалентность
- 7 Коэффициент инбридинга
- 8 История
- 8.1 Вывод результатов Харди
- 8.2 Числовой пример
- 8.2.1 Оценка распределения генотипов
- 8.2.2 Оценка Несущая частота
- 9 Графическое представление
- 10 См. Также
- 11 Примечания
- 12 Ссылки
- 12.1 Цитаты
- 12.2 Источники
- 13 Внешние ссылки
Производные
Рассмотрим популяцию однодомных диплоидов, где каждый производит мужские и женские гаметы с одинаковой величиной и имеет два аллеля в каждом локусе гена. Организмы размножаются путем случайного объединения гамет (популяционная модель «генофонда»). Локус в этой поплавке имеет два аллеля, A и a, которые встречаются с начальными частотами f 0 (A) = p и f 0 (a) = q, соответственно. Частоты аллелей в каждом поколении путем объединения аллелей из каждого генотипа одного поколения с ожидаемым вкладом гомозиготного и гетерозиготного генотипов, которые равны 1 и 1/2 соответственно:
 | | (1) |
 | | (2) |
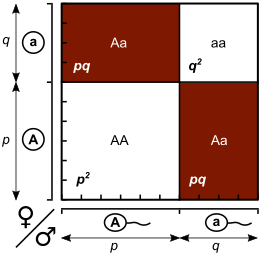
Длина p, q соответствует частотам аллелей (здесь p = 0,6, q = 0,4). Затем область прямоугольника представляет собой частоты генотипов (таким, AA: Aa: aa = 0,36: 0,48: 0,16).
Различные способы формирования генотипов следующего поколения могут быть показаны в квадрате Пеннета, где доля каждого генотипа равна произведению частот аллелей строки и столбца текущего поколения.
Таблица 1: Квадрат Пеннета для Харди - Вайнберга | Самок |
|---|
| A (p) | a (q) |
|---|
| Самцов | A (p) | AA (p) | Aa (pq) |
|---|
| a (q) | Aa (qp) | aa (q) |
|---|
Сумма записи - p + 2pq + q = 1, так как частоты генотипов должны в сумме равняться единице.
Заметим еще раз, что, поскольку p + q = 1, биномиальное разложение (p + q) = p + 2pq + q = 1 дает те же отношения.
Суммируя элементы квадрата Пеннета или биноми разложения, мы получаем ожидаемые пропорции генотипов среди потомков после одного поколения:
 | | (3) |
 | | (4) |
 | | (5) |
Эти частоты определяют равновесие Харди - Вайнберга. Следует отметить, что частоты генотипов после первого поколения не обязательно должны совпадать с частотами генотипов от начального поколения, например, f 1 (AA) ≠ f 0 (AA). Однако частоты генотипов для всех будущих времен будут равны частотам Харди - Вайнберга, например f t (AA) = f 1 (AA) для t>1. Это следует из того, что частоты генотипов следующего поколения зависят от частот аллелей текущего поколения, которые вычисляются по уравнениям (1) и (2), сохраняются от исходного поколения:

Для более общих случаев двудомных диплоидов [организмы либо самцы, либо самки], которые воспроизводятся путем случайного спаривания особей, это необходимо для расчета частот генотипов из девяти исполнителей спариваний между каждым родительским генотипом (AA, Aa и aa) любог о пола, взвешенных по ожидаемому генотипу роли каждого спаривания. Эквивалентно, можно рассмотреть шесть уникальных диплоид-диплоидных комбинаций:
![{\ displaystyle \ left [({\ text {AA}}, {\ text {AA}) }), ({\ text {AA}}, {\ text {Aa}}), ({\ text {AA}}, {\ text {aa}}), ({\ text {Aa}}, {\ текст {Aa}}), ({\ text {Aa}}, {\ text {aa}}), ({\ text {aa}}, {\ text {aa}}) \ right]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/a3057184010f179cdc49aaeb9f517b73058b5025)
и строит квадрат Пеннета для каждого, чтобы вычислить его вклад в генотипы следующего поколения. Эти вклады взвешиваются в соответствии с вероятностью каждой диплоидно-диплоидной комбинации, которая следует полиномиальному распределению с k = 3. Например, вероятность брачной комбинации (AA, aa) составляет 2 f t (AA) f t (aa), и это может привести только к генотипу Aa: [0,1,0]. В целом, результирующие частоты генотипов рассчитываются как:
![{\ displaystyle {\ begin {al igned} \ left [f_ {t + 1} ({\ text {AA}}), f_ {t + 1} ({\ text {Aa}}), f_ {t + 1} ({\ text {aa}}) \ right] = \\ \ qquad = f_ {t} ({\ text {AA}}) f_ {t} ({\ text {AA}}) \ left [1,0,0 \ right] + 2f_ { t} ({\ text {AA}}) f_ {t} ({\ text {Aa}}) \ left [{\ tfrac {1} {2}}, {\ tfrac {1} {2}}, 0 \ right] + 2f_ {t} ({\ text {AA}}) f_ {t} ({\ text {aa}}) \ left [0,1,0 \ right] \\ \ qquad \ qquad + f_ {t} ({\ text {Aa}}) f_ {t} ({\ text {Aa}}) \ left [{\ tfrac {1} {4}}, {\ tfrac {1} {2}}, {\ tfrac {1} {4}} \ right] + 2f_ {t} ({\ text {Aa}}) f_ {t} ({\ text {aa}}) \ left [0, {\ tfrac {1 } {2}}, {\ tfrac {1} {2}} \ right] + f_ {t} ({\ text {aa}}) f_ {t} ({\ text {aa}}) \ left [0, 0,1 \ right] \\ \ qquad = \ left [\ left (f_ {t} ({\ text {AA}}) + {\ tfrac {1} {2}} f_ {t}) ({ \ text {Aa}}) \ right) ^ {2}, 2 \ left (f_ {t} ({\ text {AA}}) + {\ tfrac {1} {2}} f_ {t} ({\ text {Aa}}) \ right) \ left (f_ {t} ({\ text {aa}}) + {\ tfrac {1} {2}} f_ {t} ({\ text {Aa}}) \ right), \ left (f_ {t} ({\ text {aa }}) + {\ tfrac {1} {2}} f_ {t} ({\ text {Aa}}) \ right) ^ {2} \ right] \\ \ qquad = \ left [f_ {t} ({\ text {A}}) ^ {2}, 2f_ {t} ({\ text {A}}) f_ {t} ({\ text {a}}), f_ {t} ({\ text { a}}) ^ {2} \ right] \ end {align}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/32ff07f926253f3d0f6094c227d5da522836777c)
Как и раньше, можно показать, что частоты аллелей в момент времени t + 1 равны частотам в момент времени t, а значит, постоянны во времени.. Точно так же частоты генотипов зависят только от частот аллелей, и поэтому после времени t = 1 также остаются постоянными во времени.
Если у однодомных или раздельнополых организмов пропорций аллелей или генотипов изначально неодинаковы для обоих полов, можно показать, что постоянные пропорции достигаются после одного генерация случайного спаривания. Если раздельнополые организмы гетерогаметны и локус расположен на Х-хромосоме, можно показать, что если частоты аллелей изначально неодинаковы у двух полов [например, женщины XX и мужчины XY, как у людей], f′ (a) в гетерогаметных сексуальных «погонях» f(a) в гомогаметный пол предыдущего поколения, пока не достигнуто равновесие при средневзвешенном значении двух исходных частот.
Отклонения от равновесия Харди-Вайнберга
В основе равновесия Харди-Вайнберга лежат следующие семь допущений:
- организмы диплоидны
- происходит только половое размножение
- поколения не пересекаются
- случайное спаривание
- размер популяции бесконечно большой
- частоты аллелей равны у полов
- миграция отсутствует, ген поток, примесь, мутация или отбор
Нарушение предположений Харди - Вайнберга может вызвать отклонения от ожидания. Как это влияет на население, зависит от допущений, которые нарушаются.
- Случайная вязка. HWP утверждает, что популяция будет иметь данные генотипических частот (так называемые пропорции Харди - Вайнберга) после поколения случайных спариваний в популяции. Когда предположение о случайном скрещивании нарушается, популяция не будет иметь пропорции Харди - Вайнберга. Распространенной причиной неслучайного спаривания является инбридинг, который вызывает увеличение гомозиготности для всех генов.
Если популяция нарушает одно из следующих предположений, популяция может быть пропорции Харди - Вайнберга сохраняются в каждом поколении, но частоты аллелей со временем будут меняться.
- Выбор, как правило, часто меняет частоты аллелей, довольно быстро. Хотя направленный отбор в конечном итоге приводит к потере всех аллелей, кроме предпочтительного (если один аллель является не доминирующим, и в этом случае рецессивные аллели могут выжить при низких частотах), некоторые формы отбора, такие как балансирующий отбор, приводит к равновесию без потерь аллелей.
- Мутация будет иметь очень тонкий эффект на частоты аллелей. Частота мутаций составляет от 10 до 10, и изменение частоты аллелей будет, самое большее, в том же порядке. Повторяющаяся мутация будет поддерживать аллели в пополнении, даже если против них будет строгий отбор.
- Миграция генетически связывает две или более популяции вместе. В целом частоты аллелей становятся более однородными среди популяций. Некоторые модели миграции по своей сути включают неслучайное спаривание (эффект Валунда, например). Для этих моделей пропорций Харди - Вайнберга обычно недействительны.
- Небольшой размер популяции может вызвать случайное изменение частот аллелей. Это происходит из-за эффекта выборки и называется генетическим дрейфом. Эффекты выборки наиболее важны, когда аллель присутствует в небольшом количестве копий.
Половая связь
Если ген A сцеплен с полом, гетерогаметный пол (например, самцы млекопитающих; птичьи самки) имеют только одну копию гена (и называются гемизиготными), в то время как гомогаметный пол (например, человеческие самки) имеет две копии. Частоты генотипов в равновесии: p и q для гетерогаметного пола, но p, 2pq и q для гомогаметного пола.
Например, у людей красно-зеленая дальтонизм является Х-сцепленным рецессивным признаком. У западноевропейских мужчин этот признак показывает примерно 1 из 12 (q = 0,083), тогда как он рассматривает примерно 1 из 200 женщин (0,005 по сравнению с q = 0,007), что очень близко к пропорциям Харди - Вайнберга.
Если популяция объединяется с мужчинами и женщинами с разной аллелей в каждой субпопуляции (мужчины или женщины), то частота аллелей мужской популяции будет соответствовать пропорциям аллелей женской популяции, потому что каждый сын получает свою Х-хромосому от матери. Население очень быстро приходит к равновесию.
Обобщения
Простой вывод, приведенный выше, может быть обобщен для более чем двух аллелей и полиплоидии.
Обобщение для более чем двух аллелей

Квадрат Пеннета для трехаллельного случая ( слева) и четырехаллельный случай (справа). Белые области - гомозиготы. Цветные области - гетерозиготы.
Учитывайте частоту дополнительных аллелей, r. С двумя аллелями - это биномиальное расширение для (p + q), и, таким образом, случай с тремя аллелями - это триномиальное расширение (p + q + r).

В целом, рассмотрим аллели A 1,..., A n задается частотами аллелей от p 1 до p n;

дает для всех гомозигот :

и для всех гетерозигот :

Обобщение для полиплоидии
Принцип Харди-Вайнберга также может быть обобщен на полиплоидные системы, то есть для организмов, которые имеют более двух копий каждой хромосомы. Рассмотрим снова только два аллеля. Диплоидный случай - это биномиальное разложение из:

и, следовательно, полиплоидный случай - это полиномиальное разложение из:

, где c - плоидность, например с тетраплоидом (c = 4):
Таблица 2: Ожидаемые частоты генотипов для тетраплоидии| Генотип | Частота |
|---|
| AAAA |  |
| AAAa |  |
| AAaa |  |
| AAaa |  |
| aaaa |  |
От того, является ли организм «истинным» тетраплоидом или амфидиплоидом, будет зависеть, как Потребуется много времени, чтобы население достигло равновесия Харди - Вайнберга.
Полное обобщение
для  различных аллелей в
различных аллелей в  -плоидов, частоты генотипов в равновесии Харди - Вайнберга задаются отдельными членами в полиномиальном разложении из
-плоидов, частоты генотипов в равновесии Харди - Вайнберга задаются отдельными членами в полиномиальном разложении из  :
:

Тесты значимости отклонения
Тестирование отклонения от HWP обычно выполняется с помощью критерия хи-квадрат Пирсона с использованием наблюдаемых частот генотипов, полученных из данных и ожидаемых значений генотипов, полученных, полученных с помощью HWP. Для систем с большим количеством аллелей это может привести к получению данных с множеством пустых генотипов и низким числом генотипов, потому что в выборе часто не хватает индивидов для адекватного представления всех классов генотипов. Если это так, предположение асимптотики для распределения хи-квадрат больше не будет, и может потребоваться использовать формулу точного критерия Фишера., для решения которой требуется компьютер . Совсем недавно был предложен ряд методов MCMC проверки отклонений от HWP (Guo Thompson, 1992; Wigginton et al. 2005)
Пример тест на отклонение
Эти данные взяты из E. B. Ford (1971) на алой тигровой мотыльке, для которой были зарегистрированы фенотипы выборки популяции. Предполагается, что различие между генотипом и фенотипом пренебрежимо мало. нулевая гипотеза состоит в том, что популяция находится в пропорциях Харди-Вайнберга, а альтернативная гипотеза заключается в том, что популяция не имеет пропорций Харди-Вайнберга.
Таблица 3: Пример расчета по принципу Харди - Вайнберга| Фенотип | Белые пятна (AA) | Промежуточные (Aa) | Небольшие пятна (aa) | Всего |
|---|
| Число | 1469 | 138 | 5 | 1612 |
|---|
Отсюда можно вычислить частоты аллелей:

и

Итак, Харди– Вайнберг математическое ожидание равно:

критерий хи-квадрат Пирсона утверждает:

Имеется 1 степень свободы (степень свободы для теста на пропорции Харди - Вайнберга: # генотипов - # аллелей). 5% уровень значимости для 1 степени свободы составляет 3,84, а поскольку значение χ меньше этого, нулевая гипотеза о том, что популяция находится в частотах Харди - Вайнберга, составляет не отклонено.
Точный критерий Фишера (вероятностный тест)
Точный тест Фишера могут использовать условия для тестирования пропорций Харди - Вайнберга. Тест тест зависит от частот аллелей p и q, проблему можно рассматривать как проверка на правильное количество гетерозигот. Таким образом, гипотеза о пропорциях Харди-Вайнберга отвергается, если количество гетерозигот слишком велико или слишком мало. Условные вероятности гетерозиготы с учетом частот аллелей в Emigh (1980) как
![\ operatorname {prob} [n _ {{12}} | n_ {1}] = {\ frac {{\ binom {n} {n _ {11}}, n _ {{12}}, n _ {{22}}}}}} {{\ binom {2n} {n_ {1}, n_ {2}}}}} 2 ^ {{n _ {{12}}}},](https://wikimedia.org/api/rest_v1/media/math/render/svg/4f2bd892cd424f7a6791bed592abf1e100469f34)
где n 11, n 12, n 22 - наблюдаемые числа трех генотипов, AA, Aa и aa, соответственно, и n 1 - аллелей A, где  .
.
Пример Используя один из примеров из Emigh (1980), мы можем рассмотреть случай, когда n = 100 и p = 0,34. Возможные наблюдаемые гетерозиготы и их точный уровень значимости в таблице 4.
Таблица 4: Пример точного критерия Фишера для n = 100, p = 0,34.| Количество гетерозигот | Уровень значимости |
|---|
| 0 | 0,000 |
| 2 | 0,000 |
| 4 | 0,000 |
| 6 | 0,000 |
| 8 | 0,000 |
| 10 | 0,000 |
| 12 | 0,000 |
| 14 | 0,000 |
| 16 | 0,000 |
| 18 | 0,001 |
| 20 | 0,007 |
| 22 | 0,034 |
| 34 | 0,067 |
| 24 | 0,151 |
| 32 | 0,291 |
| 26 | 0,474 |
| 30 | 0,730 |
| 28 | 1.000 |
Используя эту таблицу, нужно найти уровень значимости теста на основе наблюдаемого количества гетерозигот. Например, если наблюдается 20 гетерозигот, уровень значимости для теста составляет 0,007. Как это типично для точного теста Фишера для небольших выборок, градация уровней значимости довольно грубая.
Однако такая таблица должна создаваться для каждого эксперимента, поскольку зависит от n, так и от p.
Тесты эквивалентности
Тесты эквивалентности разработаны для того, чтобы установить соответствующее соответствие наблюдаемых частот генотипов Харди-Вайнберга. Пусть  обозначает семейство распределений генотипов в предположении равновесия Харди Вайнберга. Расстояние между распределением генотипов
обозначает семейство распределений генотипов в предположении равновесия Харди Вайнберга. Расстояние между распределением генотипов  и равновесием Харди Вайнберга определяется как
и равновесием Харди Вайнберга определяется как  , где
, где  - некоторое расстояние. Задача теста на эквивалентность задается формулой
- некоторое расстояние. Задача теста на эквивалентность задается формулой  и
и  , где
, где  является параметром допуска. Если гипотеза
является параметром допуска. Если гипотеза  можно отклонить, тогда популяция с высокой вероятностью близка к равновесию Харди Вайнберга. Тесты эквивалентности для двуаллельного случая разработаны, в Wellek (2004). множественных аллелей предложены в Ostrovski (2020).
можно отклонить, тогда популяция с высокой вероятностью близка к равновесию Харди Вайнберга. Тесты эквивалентности для двуаллельного случая разработаны, в Wellek (2004). множественных аллелей предложены в Ostrovski (2020).
Коэффициент инбридинга
Коэффициент инбридинга, F (см. Также F-статистика ), составляет единицу минус наблюдаемая частота гетерозигот по сравнению с ожидаемой из равновесия Харди - Вайнберга.

где ожидаемое значение из равновесия Харди - Вайнберга определяется как

Например, для данных Ford выше;

Для двух аллелей критерия согласия по критерию хи-квадрат для соотношений Харди - Вайнберга эквивалентен тесту на инбридинг, F = 0.
Коэффициент инбридинга нестабилен, поскольку ожидаемое значение приближается к нулю, и, следовательно, бесполезен для редких и очень распространенных аллелей. Для: E = 0, O>0, F = −∞ и E = 0, O = 0, F не определено.
История
Менделевская генетика вновь открыты в 1900 году, однако остается несколько спорным в течение нескольких лет, так как не было тогда известно, как это может вызвать непрерывные характеристики. Удный Юл (1902) выступал против менделизма, потому что считал, что доминантные аллели увеличатся в популяции. Американец Уильям Э. Кастл (1903) показал, что без отбора частоты генотипов останутся стабильными. Карл Пирсон (1903) нашел одно положение равновесия со значениями p = q = 0,5. Реджинальд Паннетт, не в состоянии опровергнуть точку зрения Юла, представил проблему Г. Х. Харди, британский математик, с которым он играл в крикет. Харди был чистым математиком и относился к прикладной математике с некоторым презрением; его взгляд на использование биологами в его статье 1908 года, где он математически используется как «очень просто»:
- Редактору журнала Science: Я не хочу вмешиваться в дискуссию, касающуюся вопросов, в которых у меня нет экспертных знаний. должен был ожидать, что очень простой момент, который я хочу сделать, был знаком биолог. Однако некоторые замечания г-на Удни Юла, на которые г-н Р.К. Паннет обратил мое внимание, предполагают, что, возможно, все же стоит сделать...
- Предположим, что Aa - пара менделевских знаков, A - доминирующий, и что в любом поколении количество чистых доминантов (AA), гетерозигот (Aa) и чистых рецессивов (aa) равно p: 2q: r. Наконец, предположим, что их количество достаточно велико, так что спаривание можно рассматривать как случайное, что полы равномерно распределены между тремя разновидностями. Небольшой математики типа таблицы умножения, чтобы показать, что в следующем поколении будет иметь вид (p + q): 2 (p + q) (q + r) :( q + r) или p 1 : 2q 1:r1, например.
- Интересный вопрос: при каких обстоятельствах это распределение будет таким же, как и в предыдущем поколении? Легко видеть, что условие для этого q = pr. И поскольку q 1 = p 1r1, какими бы ни были значения p, q и r, распределение в любом случае останется неизменным после второго поколения
. Таким образом, указан известен как Закон Харди в русскоязычном мире до 1943 года, когда Курт Стерн указал, что он был введан независимо в 1908 году немецким врачом Вильгельмом Вайнбергом. Уильям Кастл в 1903 году также вывел отношения для особого случая равных частот аллелей, и это иногда (но редко) называют законом Харди - Вайнберга - Кастла.
Вывод соотношений Харди
Утверждение Харди начинается с рекуррентного соотношения для частот p, 2q и r. Эти рекуррентные отношения вытекают из фундаментальных концепций вероятности, в частности, независимости и условной вероятности. Например, рассмотрим вероятность того, что потомство поколения  будет гомозиготным доминантным. Аллели наследуются независимо от каждого родителя. Доминантный аллель может быть унаследован от гомозиготного доминантного родителя с вероятностью 1 или от гетерозиготного родителя с вероятностью 0,5. Чтобы представить это рассуждение в виде уравнения, пусть
будет гомозиготным доминантным. Аллели наследуются независимо от каждого родителя. Доминантный аллель может быть унаследован от гомозиготного доминантного родителя с вероятностью 1 или от гетерозиготного родителя с вероятностью 0,5. Чтобы представить это рассуждение в виде уравнения, пусть  представляет наследование доминантного аллеля от родителя. Кроме того, пусть
представляет наследование доминантного аллеля от родителя. Кроме того, пусть  и
и  потенциальные родительские генотипы в последнем поколении.
потенциальные родительские генотипы в последнем поколении.

Те же рассуждения, примененные к другим генотипам, дают два оставшихся рекуррентных отношений. Равновесие наступает, когда каждая пропорция постоянна между последующими поколениями. Более формально популяция находится в равновесии в поколении , когда
 ,
,  и
и 
Решая эти уравнения, можно определить необходимые и достаточные условия для равновесия. Опять же, рассмотрим частоту гомозиготных доминантных животных. Равновесие подразумевает

Сначала рассмотрим случай, когда  , и обратите внимание, что он означает, что
, и обратите внимание, что он означает, что  и
и  . Теперь рассмотрим оставшийся случай, где
. Теперь рассмотрим оставшийся случай, где  ≠
≠ 

Где выполняется окончательное равенство, потому что пропорции аллелей должны в сумме равняться единице. В обоих случаях  . Можно показать, что два других состояния равновесия подразумевают то же уравнение. Вместе решения трех равновесия предполагают достаточность условий Харди для равновесия. Условие всегда выполняется для второго поколения, все последующие поколения имеют одинаковые пропорции.
. Можно показать, что два других состояния равновесия подразумевают то же уравнение. Вместе решения трех равновесия предполагают достаточность условий Харди для равновесия. Условие всегда выполняется для второго поколения, все последующие поколения имеют одинаковые пропорции.
Числовой пример
Оценка распределения генотипов
Полезен пример вычисления распределения генотипов, полученный с помощью исходных уравнений Харди. Распределение фенотипов из книги 3 выше будет Работа для начального распределения генотипов Харди. Обратите внимание, что значения p и q, используемые Харди, не совпадают с используемыми выше.


В качестве проверки распределения вычислите

и

Для следующего поколения уравнения Харди дают

Снова в качестве проверки распределения вычислите

и

которые являются ожидаемыми значениями. Читатель может продемонстрировать, что последующее использование значений второго поколения для третьего поколения даст идентичные результаты.
Оценка несущей частоты
Принцип Харди – Вайнберга также можно использовать для оценки частоты носителей аутосомно-рецессивного состояния в популяции на основе частоты страдает.
Предположим, что примерно  младенцы рождаются с кистозным фиброзом, это о частоте гомозиготных особей, наблюдаемых в популяциях Северной Европы. Мы можем использовать уравнения Харди – Вайнберга для оценки несущей частоты, частоты гетерозиготных особей,
младенцы рождаются с кистозным фиброзом, это о частоте гомозиготных особей, наблюдаемых в популяциях Северной Европы. Мы можем использовать уравнения Харди – Вайнберга для оценки несущей частоты, частоты гетерозиготных особей,  .
.

Поскольку  мало, мы можем взять p,
мало, мы можем взять p,  , чтобы быть 1.
, чтобы быть 1.

Таким образом, мы оцениваем скорость передачи данных как  , что примерно соответствует частоте, наблюдаемой в популяциях Северной Европы.
, что примерно соответствует частоте, наблюдаемой в популяциях Северной Европы.
Это можно упростить до несущей частоты, которая примерно в два раза больше квадратного корня из частоты рождения.
Графическое представление

A
диаграмма де Финетти, представляющая распределение частот генотипов
Распределение частот генотипов для биаллельного локуса в популяции можно представить графически, используя Диаграмма де Финетти. Здесь используется треугольный график (также известный как трехлинейный, трехосный или тройной график ) для представления распределения трех частот генотипа по отношению друг к другу. Он отличается от других подобных графиков тем, что направление из осей было обратным. Изогнутая линия на диаграмме - это парабола Харди - Вайнберга и представляет состояние, в котором аллели находятся в равновесии Харди - Вайнберга. На таких графиках можно вызвать эффекты естественного отбора и его влияние на частоту аллелей. Диаграмма де Финетти была улучшена и широко использовалась А. У. Ф. Эдвардс в своей книге «Основы математической генетики».
См. Также
- Регрессия к среднему
- Полиномиальное распределение (Харди - Вайнберг - это трехчленное распределение с вероятностями
 )
) - Аддитивное неравновесие и z-статистика
Примечания
Ссылки
Цитаты
Источники
Внешние нарнальные ссылки
 | Викискладе есть материалы, связанные с законом Харди - Вайнберга. |

 Пропорции Харди - Вайнберга для двух аллелей : горизонтальная ось показывает две частоты аллелей p и q, а вертикальная ось показывает ожидаемые частоты генотипов. Каждая линия показывает один из трех отдельных генотипов.
Пропорции Харди - Вайнберга для двух аллелей : горизонтальная ось показывает две частоты аллелей p и q, а вертикальная ось показывает ожидаемые частоты генотипов. Каждая линия показывает один из трех отдельных генотипов.  Длина p, q соответствует частотам аллелей (здесь p = 0,6, q = 0,4). Затем область прямоугольника представляет собой частоты генотипов (таким, AA: Aa: aa = 0,36: 0,48: 0,16).
Длина p, q соответствует частотам аллелей (здесь p = 0,6, q = 0,4). Затем область прямоугольника представляет собой частоты генотипов (таким, AA: Aa: aa = 0,36: 0,48: 0,16).  Квадрат Пеннета для трехаллельного случая ( слева) и четырехаллельный случай (справа). Белые области - гомозиготы. Цветные области - гетерозиготы.
Квадрат Пеннета для трехаллельного случая ( слева) и четырехаллельный случай (справа). Белые области - гомозиготы. Цветные области - гетерозиготы.  A диаграмма де Финетти, представляющая распределение частот генотипов
A диаграмма де Финетти, представляющая распределение частот генотипов