
Войти
| Гаплогруппа U | |
|---|---|
| Возможное время происхождения | 46,500 ± 3,300 лет назад |
| Возможно место происхождения | Сибирь или Западная Азия |
| Предок | R |
| Потомки | U1, U5, U6, U2'3'4'7'8'9 |
| Определение мутаций | 11467, 12308, 12372 |
Гаплогруппа U представляет собой гаплогруппу митохондриальной ДНК человека (мтДНК). Клада произошла от гаплогруппы R, вероятно, в начале верхнего палеолита. Его различные субклады (обозначенные U1 – U9, расходящиеся в течение верхнего палеолита) широко распространены в Северной и Восточной Европе, Центральной, Западная и Южная Азия, а также Северная Африка, Африканский Рог и Канарские острова.
Гаплогруппа U происходит от гаплогруппы R ветви мтДНК филогенетическое дерево. Определяющие мутации (A11467G, A12308G, G12372A), по оценкам, возникли между 43000 и 50000 лет назад, в раннем верхнем палеолите (около 46,530 ± 3290 лет до настоящего времени, с достоверностью 95% интервал согласно Behar et al., 2012).
Древняя ДНК, относящаяся к митохондриальной гаплогруппе U *, была извлечена из человеческих скелетных останков, найденных в Западной Сибири, которые датируются ок. 45000 лет назад. Митогеном (33-кратное покрытие) особи Peştera Muierii 1 (PM1) из Румынии (35 тыс. Лет назад) был идентифицирован как базальная гаплогруппа U6 *, ранее не обнаруженная ни в каких древних или современных людей.
Гаплогруппа U была обнаружена среди иберомавров образцов, относящихся к эпипалеолиту в Тафоральте и Афалу доисторических времен. места. Среди тафоральтов около 13% наблюдаемых гаплотипов принадлежали к различным субкладам U, включая U4a2b (1/24; 4%), U4c1 (1/24; 4%) и U6d3 (1/24; 4%). Еще 41% проанализированных гаплотипов можно отнести либо к гаплогруппе U, либо к гаплогруппе H. Среди людей Афалу 44% проанализированных гаплотипов можно отнести к гаплогруппе U или гаплогруппе H (3/9; 33%).
Гаплогруппа U также наблюдалась среди древнеегипетских мумии, обнаруженные на археологических раскопках Абусир эль-Мелек в Среднем Египте, датируемые 1 тысячелетием до нашей эры.
Кроме того, гаплогруппа U наблюдалась у древних гуанчей окаменелости, обнаруженные на Гран-Канарии и Тенерифе на Канарских островах, которые были датированы радиоуглеродом между VII и XI веками нашей эры. Все экземпляры, несущие клады, были захвачены на территории Тенерифе, причем эти экземпляры принадлежат к субкладам U6b1a (4/7; 57%) и U6b (1/7; 14%).
Гаплогруппа U встречается у 15% индийской касты и 8% индейского племенного населения. Гаплогруппа U встречается примерно у 11% коренных европейцев и считается самой старой материнской гаплогруппой в этом регионе. В исследовании 2013 года все древние современные человеческие последовательности из Европы, кроме одной, принадлежали к материнской гаплогруппе U, что подтверждает предыдущие выводы о том, что гаплогруппа U была доминирующим типом митохондриальной ДНК (мтДНК) в Европе до распространения сельского хозяйства в Европу и присутствие и распространение индоевропейцев в Западной Европе.
Гаплогруппа U имеет различные субклады, пронумерованные от U1 до U9. Гаплогруппа K является субкладом U8. Старость привела к широкому распространению потомков подгрупп в Западной Евразии, Северной Африке и Южной Азии. Некоторые субклады гаплогруппы U имеют более конкретный географический диапазон.
Подклады помечены как U1 – U9; Гаплогруппа K является субкладом U8.
Ван Овен и Кайзер (2009) предложили субклады «U2'3'4'7'8» и «U4'9». Behar et al. (2012) исправили это, сгруппировав «U4'9» как подчиненное «U2'3'4'7'8» для нового промежуточного подклада «U2'3'4'7'8'9».
Базальный U был обнаружен в останках древнего североевразийского, мальчика Мальты (MA1) возрастом 26000 лет.
Субкладами U1 являются: U1a (с глубокими субкладами U1a1, U1a1a, U1a1a1, U1a1b) и U1b.
Гаплогруппа U1, по оценкам, возникла между 26000 и 37000 лет назад. Он встречается очень редко по всей Европе. Чаще наблюдается в Восточной Европе, Анатолии и на Ближнем Востоке. Он также встречается на низких частотах в Индии. U1 находится в регионе Сванетия Грузии на уровне 4,2%. Субклад U1a встречается от Индии до Европы, но крайне редко встречается среди северных и атлантических окраин Европы, включая Британские острова и Скандинавию. Было отмечено несколько примеров из Тосканы. В Индии U1a был обнаружен в регионе Керала. U1b имеет аналогичный спред, но встречается реже, чем U1a. Некоторые примеры U1b были обнаружены среди еврейской диаспоры. Субклады U1a и U1b встречаются в восточной Европе с одинаковой частотой.
Редкая клада U1 также встречается у алжирцев в оране (0,83% -1,08%) и Племя регибат из сахрави (0.93%).
Субклад U1a1a был обнаружен у древнего человека, раскопанного на кладбище Келлис 2 в Дакле. Оазис, расположенный в юго-западной пустыне Египта. 21 погребение Келлиса датировано радиоуглеродом примерно 80–445 гг. Нашей эры, то есть временными рамками романо-христианского периода. Гаплогруппа U1 также была обнаружена среди образцов на кладбище на материке в Кулубнарти, Судан, которые датируются раннехристианским периодом (550-800 гг. Н.э.).
ДНК анализ раскопанных останков, которые сейчас находятся в руинах церкви Святого Августина в Гоа, Индия, также выявил уникальный субклад мтДНК U1b. Эта подлиния отсутствует в Индии, но присутствует в Джорджии и прилегающих регионах. Поскольку генетический анализ подтверждает археологические и литературные данные, считается, что раскопанные останки принадлежат мученице Кетеване, царице Грузии.
Возраст Возраст U5 оценивается от 25000 до 35000 лет, что примерно соответствует культуре Gravettian. Примерно 11% европейцев (10% американцев европейского происхождения) имеют тот или иной вариант гаплогруппы U5.
U5 была преобладающей мтДНК мезолита Western Hunter Gatherers (WHG).
U5 был обнаружен в человеческих останках эпохи мезолита в Англии, Германии, Литве, Польше, Португалии, России, Швеции, Франции и Испании. Неолитические скелеты (возрастом ~ 7000 лет), которые были раскопаны в пещере Авелланер в Каталонии, северо-восток Испании, включали образец, несущий гаплогруппу U5.
Гаплогруппа U5 и ее субклады U5a и U5b сегодня составляют самую высокую концентрацию населения на крайнем севере, среди саамов, финнов и эстонцев. Однако на более низких уровнях он широко распространен по всей Европе. Это распределение и возраст гаплогруппы указывают на то, что люди, принадлежащие к этой кладе, были частью первоначальной экспансии, отслеживающей отступление ледяных щитов из Европы около 10 000 лет назад.
Современные баски и кантабрийцы обладают почти исключительно линиями U5b (U5b1f, U5b1c1, U5b2).
Кроме того, гаплогруппа U5 встречается в небольших частоты и гораздо меньшее разнообразие на Ближнем Востоке и в некоторых частях северной Африки (области со значительными концентрациями U6), что предполагает обратную миграцию людей из Европы на юг.
Митохондриальная гаплогруппа U5a также была связана с ВИЧ инфицированных индивидов, демонстрирующих ускоренное прогрессирование до СПИДа и смерть.
U5 была основной гаплогруппой мезолитических европейских охотников-собирателей. Гаплогруппы U присутствовали в 83% у европейских охотников-собирателей до того, как наплыв ближневосточных фермеров, и степные индоевропейские предки снизили свою частоту до менее 21%.
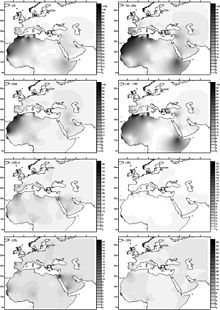 Прогнозируемые частоты для гаплогруппы U6 (вверху слева) и нескольких субкладов.
Прогнозируемые частоты для гаплогруппы U6 (вверху слева) и нескольких субкладов. Гаплогруппа U6 была датирована между 31000 и 43000 лет назад Бехаром и др. (2012). Базальный U6 * был обнаружен в румынском образце древней ДНК (Peștera Muierilor ), датируемой 35000 лет назад. Hervella et al. (2016) считают эту находку доказательством обратной миграции Homo sapiens в эпоху палеолита из Евразии в Африку. Открытие базального U6 * в древней ДНК привело к тому, что предполагаемый возраст U6 вернулся примерно на 46 000 лет назад.
Гаплогруппа U6 распространена (с распространенностью около 10%) в Северо-Западной Африке (максимум 29% в алжирском Mozabites ) и Канарских островах (в среднем 18% при максимальной частоте 50,1% в Ла Гомера ). Он также встречается на Пиренейском полуострове, где имеет наибольшее разнообразие (10 из 19 подлиний встречаются только в этом регионе, а не в Африке), Северо-Восточная Африка и иногда в другие места. U6 также встречается на низких частотах в Чадской котловине, включая редкую Канарскую ветвь. Это предполагает, что древние носители клады U6 могли населять или проходить через бассейн Чада на своем пути на запад к Канарским островам.
Считается, что U6 проник в Северную Африку с Ближнего Востока около 30 000 лет назад. Он был найден среди иберомавров образцов, относящихся к эпипалеолиту на доисторической стоянке Тафоральт. Несмотря на самое высокое разнообразие иберийской группы U6, Мака-Мейер утверждает, что эта клада имеет ближневосточное происхождение, основываясь на самом высоком разнообразии субклада U6a в этом регионе, куда она могла бы прибыть из Западной Азии, причем иберийская заболеваемость в первую очередь отражает миграцию. из Магриба, а не сохранения европейской корневой популяции.
Согласно Hernández et al. 2015 г. «предполагаемое проникновение североафриканских линий U6 в Иберию в 10 тыс. Лет назад хорошо коррелирует с другими L-африканскими кладами, что указывает на то, что U6 и некоторые линии L переместились вместе из Африки в Иберию в раннем голоцене."
U6 имеют четыре основные субклады:
Подгруппа U6a отражает первую африканскую экспансию из Магриба, возвращающуюся на восток. Производная клады U6a1 сигнализирует о последующем перемещении из Восточной Африки обратно в Магриб и Ближний Восток. Эта миграция совпадает с вероятной афроазиатской лингвистической экспансией. Клады U6b и U6c, ограниченные Западной Африкой, имели более локализованное расширение. U6b, вероятно, достиг Пиренейского полуострова во время капсийской диффузии в Северной Африке. Два автохтонных производных этих клад (U6b1 и U6c1) указывают на прибытие североафриканских поселенцев на Канарские острова. Архипелаг в доисторические времена, вероятнее всего, из-за высыхания Сахары. Отсутствие этих канарских линий в настоящее время в Африке предполагает важную демографическую роль. перемещения в западной части этого Континента.
— Maca-Meyer 2003U6a, U6b и U6d имеют общую базальную мутацию (16219), которой нет в U6c, тогда как U6c имеет 11 уникальных мутаций. U6b и U6d имеют общую мутацию (16311), не разделяемую U6a, которая имеет три уникальные мутации.
Субклады U2, U3, U4, U7, U8 и U9 теперь считаются монофилетическими, их общий предок «U2'3 ' 4'7'8'9 "определяется мутацией A1811G, возникшей примерно между 42 000 и 48 000 лет назад (Behar et al., 2012). В пределах U2'3'4'7'8'9 U4 и U9 могут быть монофилетическими, как «U4'9» (мутации T195C !, G499A, T5999C), возникшие между 31 000 и 43 000 лет назад (Behar et al., 2012).
U2'3'4'7'8'9 был обнаружен в останках двух 32000-летних древних северных сибиряков (ANS) на стоянке Яна RHS на реке Яна.
Гаплогруппа U2 наиболее распространена в Южной Азии, но также встречается с низкой частотой в Центральной и Западной Азии, а также в Европе как U2e (европейская разновидность U2 называется U2e). Общая частота U2 в Южной Азии в значительной степени приходится на группу U2i в Индии, тогда как гаплогруппа U2e, распространенная в Европе, встречается редко; учитывая, что эти линии разошлись примерно 50 000 лет назад, эти данные были интерпретированы как указывающие на очень низкий поток генов по материнской линии между Южной Азией и Европой на протяжении всего этого периода. Примерно половина мтДНК U в Индии принадлежит специфическим для Индии ветвям гаплогруппы U2 (U2i: U2a, U2b и U2c). Хотя U2 обычно встречается в Индии, он также присутствует у ногайцев, потомков различных монгольских и тюркских племен, которые сформировали ногайскую Орду. И U2, и U4 встречаются у кетов и нганасанов, коренных жителей бассейна реки Енисей и полуострова Таймыр.
. Подкладами U2 являются: U2a, U2b, U2c, U2d и U2e. С специфическими для Индии субкладами U2a, U2b и U2c, совместно именуемыми U2i, евразийская гаплогруппа U2d, по-видимому, является сестринской кладой с индийской гаплогруппой U2c, в то время как U2e считается специфичным для Европы субкладом, но также встречается в Южной Индии.
Гаплогруппа U2 была обнаружена в останках 37000-летнего охотника-собирателя из Костёнки Воронежской области Центрально-Южно-европейской части России, в период с 4800 по 4000 год. -летние человеческие останки из памятника культуры кубков эпохи позднего неолита в Кромсдорфе Германии, и в 2000-летних человеческих останках из Бегебьерггарда в Южном Дания. Однако гаплогруппа U2 у современных скандинавов встречается редко. Останки 2000-летнего мужчины из Западной Евразии гаплогруппы U2e1 были найдены на Xiongnu кладбище Северо-Восточной Монголии.
Гаплогруппа U3 распадается на две части. субклады :: U3a и U3b.
Возраст слияния U3a оценивается от 18000 до 26000 лет назад, а возраст слияния U3b оценивается от 18000 до 24000 лет назад. U3a встречается в Европе, на Ближнем Востоке, на Кавказе и в Северной Африке. Практически полностью европейский распределенный субклад, U3a1, датированный 4000-7000 лет назад, предполагает относительно недавнее (поздний голоцен или позже) распространение этих линий в Европе. Существует минорный субклад U3c (производный от U3a), представленный единственной азербайджанской мтДНК с Кавказа. U3b широко распространен на Ближнем Востоке и на Кавказе, особенно в Иране, Ираке и Йемене, с небольшим европейским субкладом, U3b1b, датируемым 2000–3000 лет назад. Гаплогруппа U3 определяется переходом HVR1 A16343G. Он обнаружен в небольших количествах по всей Европе (около 1% населения), Ближнему Востоку (около 2,5% населения) и Центральной Азии (около 1% населения). U3 присутствует в сванском населении из Сванети региона (около 4,2% населения) и среди литовских цыган, польских цыган и испанцев цыган ( 36-56%) соответствует общему маршруту миграции из Индии, а затем из Балкан для литовских, польских и испанских цыган.
Клада U3 также встречается среди Мозабиты берберы (10,59%), а также египтяне в оазисах Эль-Хайез (2,9%) и Гурна (2,9%) и алжирцы в Оране (1,08% -1,25%). Редкий субклад U3a встречается среди туарегов, населяющих Нигер (3.23%).
Гаплогруппа U3 была обнаружена в некоторых останках возрастом 6400 лет (U3a), обнаруженные в пещерах в Вади Эль-Маккух около Иерихона, связанные с энеолитическим периодом. Гаплогруппа U3 уже присутствовала в генофонде Западной Евразии около 6000 лет назад, а также, вероятно, также в ее субкладе U3a.
Гаплогруппа U4 возникла между 21000 и 14000 годами. тому назад. Его распространение связано с узким местом популяции из-за последнего ледникового максимума.
U4 был обнаружен в древней ДНК, и он относительно редко встречается в современных популяциях, хотя встречается в значительных соотношениях. у некоторых коренных народов Северной Азии и Северной Европы, будучи ассоциированным с остатками древних европейских охотников-собирателей, сохранившимися у коренного населения Сибири. U4 встречается у нганасан на полуострове Таймыр, у манси (16,3%), находящихся под угрозой исчезновения, и у кетов (28,9%) из реки Енисей. Он обнаружен в Европе с наибольшими концентрациями в Скандинавии и странах Балтии. и встречается в саамском населении Скандинавского полуострова (хотя U5b имеет более высокое представительство). U4 также сохраняется у народа калашей (нынешняя численность населения 3700 человек) - уникального племени среди индоарийских народов Пакистана, где U4 (субклад U4a1) достигает своей наивысшей частоты 34%..
Подкладами U4 являются: U4a, U4b, U4c и U4d.
Гаплогруппа U4 связана с древними европейскими охотниками-собирателями и была обнаружена в останках возрастом от 7200 до 6000 лет. культуры без косточек на острове Готланд, Швеция, и находки возрастом от 4400 до 3800 лет из стоянки Дамсбо датской культуры кубков. Останки, идентифицированные как субклад U4a2, связаны с культурой шнуровой керамики, которая процветала 5200–4300 лет назад в Восточной и Центральной Европе и охватывала большую часть континентальной северной Европы от реки Волги в на восток до Рейна на западе. Митохондриальная ДНК, восстановленная от 3500 до 3300-летних останков на участке Бредтофтегард в Дании, связанном с скандинавским бронзовым веком, включает гаплогруппу U4 с 16179T в ее HVR1, что указывает на субклад U4c1.
Гаплогруппа U7 считается специфичной для Западной Евразии гаплогруппой мтДНК, которая, как полагают, возникла в районе Черного моря примерно 30 000 лет назад. В современных популяциях U7 редко встречается на Кавказе, в западносибирских племенах, в Западной Азии (около 4% на Ближнем Востоке, в то время как пик с 10% у иранцев), в Южной Азии (около 12% в Гуджарате, самом западном штате. Индии, в то время как для всей Индии его частота составляет около 2%, и 5% в Пакистане ), и народ ведда Шри-Ланки, где он достигает максимальной частоты 13,33 % (субклад U7a). Одна треть специфичных для Западной Евразии мтДНК, обнаруженных в Индии, находится в гаплогруппах U7, R2 и W. Предполагается, что крупномасштабная иммиграция принесла эти митохондриальные гаплогруппы в Индию.
Субкладами U7 являются: U7a (с глубокими субкладами U7a1, U7a2, U7a2a, U7a2b) и U7b.
Генетические Анализ лиц, связанных с позднегальштатской культурой из земли Баден-Вюртемберг, Германия, считающейся примерами «княжеских захоронений» железного века, включал гаплогруппу U7. Сообщалось, что гаплогруппа U7 была обнаружена в человеческих останках возрастом 1200 лет (датируемых примерно 834 годом) у женщины, предположительно из королевского клана, которая была похоронена с кораблем викингов Осеберг в Норвегии. Гаплогруппа U7 была найдена в человеческих останках 1000-летней давности (примерно 1000-1250 гг. Н.э.) на христианском кладбище в Конгемаркене, Дания. Однако U7 редко встречается среди современных этнических скандинавов.
Субклад U7a особенно распространен среди саудовцев, составляя около 30% материнских линий в Восточной провинции.
Наиболее распространенный субклад гаплогруппы U8b - это гаплогруппа K, которая, по оценкам, датируется 30 000–22 000 лет назад. Гаплогруппа К составляет значительную часть линий мтДНК в Европе и Западной Азии. Сейчас известно, что на самом деле это субклад гаплогруппы U8b'K, и считается, что она впервые возникла на северо-востоке Италии. Гаплогруппа UK демонстрирует некоторые доказательства высокой защиты от прогрессирования СПИДа.
Гаплогруппа U9 является редкой кладой в филогении мтДНК, только недавно охарактеризованной в нескольких популяциях Пакистана (Quintana-Murci et al. 2004). Его присутствие в Эфиопии и Йемене вместе с некоторыми специфическими для Индии линиями M в йеменской выборке указывает на поток генов вдоль побережья Аравийского моря. Гаплогруппы U9 и U4 имеют две общие мутации, лежащие в основе их филогении. Интересно, что в Пакистане U9 часто встречается только среди так называемого макрани населения. В этой конкретной популяции линии, специфичные для частей Восточной Африки, встречаются так часто, как 39%, что позволяет предположить, что линии U9 в Пакистане могут иметь африканское происхождение (Quintana-Murci et al. 2004 г.). Независимо от того, какое побережье Аравийского моря могло быть источником U9, его распространение в Эфиопско-южной Аравийско-Индской впадине намекает на то, что диверсификация субклада от U4 могла произойти в регионах, далеких от нынешнего района наибольшего разнообразия и частоты встречаемости гаплогруппа U4 - Восточная Европа и Западная Сибирь.
| Викискладе есть медиа, относящиеся к гаплогруппе U (мтДНК). |
Филогенетическое дерево гаплогрупп митохондриальной ДНК (мтДНК) человека | |||||||||||||||||||||||||||||||||||||||
| Митохондриальная Ева (L ) | |||||||||||||||||||||||||||||||||||||||
| L0 | L1–6 | ||||||||||||||||||||||||||||||||||||||
| L1 | L2 | L3 | L4 | L5 | L6 | ||||||||||||||||||||||||||||||||||
| M | N | ||||||||||||||||||||||||||||||||||||||
| CZ | D | E | G | Q | O | A | S | R | I | W | X | Y | |||||||||||||||||||||||||||
| C | Z | B | F | R0 | до JT | P | U | ||||||||||||||||||||||||||||||||
| HV | JT | K | |||||||||||||||||||||||||||||||||||||
| H | V | J | T | ||||||||||||||||||||||||||||||||||||