Гаплогруппа ДНК Y-хромосомы,
| Гаплогруппа N |
|---|
 |
| Возможное время происхождения | 36,800 [95% ДИ 34,300–39,300] лет до настоящего момента (YFull).. 44,700 или 38,300 лн в зависимости от частоты мутаций.. 41,900 [95% ДИ 40,175-43,591] лет назад |
|---|
| Возраст слияния | 21,700 [95% ДИ 19,500–23,900] ybp (YFull).. 25,313 [95% CI 21,722-28,956] ybp |
|---|
| Возможное место происхождения | Восточная Азия |
|---|
| Предок | NO |
|---|
| Определение мутаций | M231 |
|---|
| Наивысшие частоты | Нганассан 58% -94,1%, Якуты 81,8% -94,6% (Сюй 2015) harv error: нет цели: CITEREFXu2015 (help ), угры 77,8% (ханты 64,3% -89,3%, манси 76 %), удмурты 77,8%, хакасы 41% - 65%, коми 33,3% -79,5%, ненцы 75% -92,9 % (Тундровые ненцы 97,9%, лесные ненцы 98,8%), финны 51-61% (Purps 2014) harv error: нет цели: CITEREFPurps2014 (help ), тувинцы 27,2-54,5% (Харьков 2013) ошибка: нет цели: CITEREFKharkov2013 (help ), нанайцы 46,2% (20% Хеже в КНР, 44,6% нанайцев в России, 83,8% членов рода Самар в Горинском районе Хабаровского края), Архангельск русские 42,6% (Архангельск 44,3%, Пинега 40,8%), мари 41,2%, эстонский 30,6% -40% (Purps 2014) ошибка: нет цели: CITEREFPurps2014 ( помогите ), саамы 40%, чуваши 36%, буряты 34,5% (20,2%, 25,0%, 30,9%, 48,0%), коряки 33,3%, латыши 30% (Purps 2014) ошибка harv: нет цели: CITEREFPurps2014 (help ) -43,0%, татары Поволжья 27,8%, сибирские татары 27,6%, литовцы 25% (Purps 2014) ошибка: нет цели: CITEREFPurps2014 ( справка ) -40,5%, телеуты 25,0%, северные алтайцы 21,8% (18,0% -24,6%), псковцы Русские 22,7%, башкиры 17,3%, Сибе 17,1% -18,0%, мордва 12,5% (10% - 13,3%), монголы 11%, калмыки 10,4% (Торгууд 3,4%, Дербет 5,1%, Бузава 5,3%, Хошут 38,2%), маньчжур 10% ( 5,8%, 9,1%, 11,6%, 12,5%, 14,3%), белорусы 9,7%, центрально-южные русские 9,1% (Тверь 13,2%, Курск 12,5% -13,3%, Белгород 11,9%, Кострома 11,8%, Смоленск 7,0%, Воронеж 6,3 %, орловцы 5,5%), украинцы 9,0%, южные алтайцы 7,1% (4,2% -9,7%), ханьские китайцы 6,77 % (От 0% до 21,4%), Швеция 6% (0% - 15%), ульчи 5,8%, тибетцы 5,65%, казахи 5,33% (Суан 0%, Кангли 0%, Ошакты 0%, Джетыру 1,2%, Найман 1,3%, Дулат 1,6%, Аргын 2,0%, Алимулы 2,5%, Ысты 3,5%, Керей 3,6 %, Байулы 3,9%, Албан 4,3%, Конгырат 7,4%, Кыпшак 10,3%, Джалаир 10,9%, Кожа 16,7%, Сыргели 65,6%), Уйгуры 4,89% (2,8%, 4,8%, 4,99%, 6,0%, 8,6%), корейцы 4 0,41% (1,8% Сеул-Кёнги, 3,0% Тэджон, 4,0% Сеул, 4,2% Чхунчхон, 4,4% Чолла, 4,8% Кёнсан, 6,3% Канвон, 6,58% Корейский проект генома (в основном из Ульсана ), 6,9% Чеджу), японец 1,9% (0%, 0,8%, 0,9%, 1,7%, 2,5%, 4,3%, 4,8%, 6,4%) |
|---|
Гаплогруппа N (M231) представляет собой гаплогруппу ДНК Y-хромосомы, определяемую наличием маркера однонуклеотидного полиморфизма (SNP) M231.
Чаще всего встречается у мужчин, происходящих от из северной Евразии. Это также наблюдается с меньшей частотой у популяций коренных жителей других регионов, включая Балканы, Центральную Азию, Восточную Азию и Юго-Восточную Азию.
Содержание
- 1 Происхождение
- 2 Распределение
- 2,1 N * (M231)
- 2,2 N1 (CTS11499, Z4762, CTS3750)
- 2,3 N1a (F1206 / M2013 / S11466)
- 2,4 N1a1 (M46 / Page70 / Tat, L395 / M2080)
- 2,5 N1a1a (M178)
- 2,6 N1a2 (F1008 / L666)
- 2,7 N1a2a-M128
- 2,8 N1a2b (P43)
- 2,9 N1a2b1 -B478
- 2,10 N1b (F2930)
- 2,11 N2 (Y6503)
- 2,12 Древние народы
- 3 Филогения
- 3,1 Филогенетическое древо
- 3,2 Филогенетическая история
- 3,3 Генетика
- 3,4 Субклады Y-ДНК N
- 3.5 Древо остова Y-ДНК
- 4 Ссылки
- 4.1 Сноски
- 4.2 Цитированная работа
- 4.3 Дополнительная литература
- 4.4 Филогенетика
- 5 Внешние ссылки
Происхождение
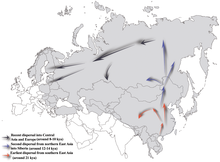
Предполагаемые доисторические маршруты миграции для линии гаплогруппы N Y-хромосомы.
Гаплогруппа NO-M214 - ее последний общий предок со своим братом, гаплогруппой O-M175 - предположительно существовало около 36 800–44 700 лет назад.
Обычно считается, что N-M231 возник в Восточной Азии примерно 19 400 (± 4800) лет назад и заселил север Евразия после последнего ледникового максимума. Самцы, несущие маркер, по-видимому, двинулись на север по мере потепления климата в голоцене, мигрируя против часовой стрелки, чтобы в конечном итоге сконцентрироваться в таких далеких областях, как Фенноскандия и Балтийский (Рутси и др., 2006). Очевидная нехватка гаплогруппы N-M231 среди коренных народов Америки указывает на то, что она распространилась после того, как Берингия была затоплена (Chiaroni, Underhill Cavalli-Sforza 2009), около 11000 лет назад.
Распространение

Прогнозируемое распределение субгаплогрупп гаплогруппы N. (A) N * -M231, (B) N1 * -LLY22g, (C) N1a-M128, (D) N1b-P43, (E) N1c-M46.
Гаплогруппа N имеет широкое географическое распространение по всей северной Евразии, а также время от времени наблюдалось в других регионах, включая Среднюю Азию и Балканы.
Наиболее часто встречается у коренных народов России, в том числе финских народов, мари, удмуртов, коми, ханты, манси, ненцы, нганасаны, тюркские народы ( Якуты, долганы, хакасы, тувинцы, татары, чуваши и др.), буряты, тунгусские народы (эвенки, эвены, негидалы, нанайцы и др.), юкагиры, луораветланцы (чукчи, коряки) и сибирские эскимосы, но некоторые субклады очень распространены в Финляндии, Эстонии, Латвии и Литве, а другие субклады встречаются с низкой частотой в Китай (И, Наси, Лхоба, Хань, китайский и т. Д.). Особенно у этнических финских и балтийских народов северной Европы, обско-угороязычных и северных самоедов западных Сибирь и тюркоязычные народы России (особенно якуты (McDonald 2005) harv error: no target: CITEREFMcDonald2005 (help ), но также алтайцы, шорцы, хакасы, чуваши, татары и башкиры ). Почти все члены гаплогруппы N среди этих популяций северной Евразии принадлежат к субкладам либо гаплогруппы N-Tat, либо гаплогруппы N-P43.
Y-хромосомы, принадлежащие N1b-F2930 / M1881 / V3743, или N1 * -CTS11499 / L735 / M2291 (xN1a-F1206 / M2013 / S11466), были обнаружены в Китае и спорадически в других частях Евразии.. N1a-F1206 / M2013 / S11466 в большом количестве встречается в Северной Евразии.
N2-Y6503, другой первичный субклад гаплогруппы N, встречается крайне редко и в основном представлен среди современных людей недавно сформированным субкладом, который фактически ограничен странами, составляющими бывшую Югославию (Босния и Герцеговина, Хорватия, Сербия и Черногория), Венгрия и Австрия. Среди других членов N2-Y6503 - венгр недавнего происхождения из Сучава в Буковине, словацкий гражданин, несколько британцев и алтайский.
N * (M231)
Y-хромосомы, которые отображают мутацию M231, которая определяет гаплогруппу N-M231, но не отображает мутации CTS11499, L735, M2291, которые определяют гаплогруппу N1, являются считается, что он принадлежит к парагруппе N-M231 *.
N-M231 * был обнаружен в небольших количествах в Китае. Из выборки 165 хань мужчин из Китая, два человека (1,2%) оказались принадлежащими к N *. (Karafet et al. 2010). Один произошел из Гуанчжоу, а другой из Сиань.
. Среди древних образцов байкальского энеолита один из образцов Шаманка II, датируемый ок. 6500 л.н., анализировался как NO1-M214.
N1 (CTS11499, Z4762, CTS3750)
В 2014 г. произошло серьезное изменение в определении субклада N1, когда LLY22g был исключен из использования в качестве основного определяющего SNP для N1 из-за сообщений о Ненадежность LLY22g. Согласно ISOGG, LLY22g проблематичен, потому что это «палиндромный маркер, который легко может быть неправильно истолкован». С тех пор название N1 было применено к кладе, отмеченной большим количеством SNP, включая CTS11499, Z4762 и CTS3750. N1 - самый недавний общий предок всех существующих членов гаплогруппы N-M231, за исключением членов редкого субклада N2-Y6503 (N2-B482). TMRCA N1 оценивается за 18 000 лет до настоящего времени (16 300–19 700 лет назад; 95% ДИ). После ревизии 2014 года положение многих примеров «N1-LLY22g» в гаплогруппе N стало неясным. Поэтому лучше проверить yfull и ISOGG 2019, чтобы понять структуру N-M231, как она понимается в настоящее время.
Однако в более ранних исследованиях сообщалось, что частота N-LLY22g достигает 30% (13/43) среди людей И из округа Бутуо, Сычуань в Юго-Западном Китае (Hammer et al. 2005, Karafet et al. 2001 и Wen2004b ошибка harvnb: нет цели: CITEREFWen2004b (справка )). Он также встречается у 34,6% людей Lhoba (Wen 2004 harvnb error: no target: CITEREFWen2004 (help ), Bo Wen 2004 ошибка harvnb: нет цели: CITEREFBo_Wen2004 (справка )). N1-LLY22g * был обнаружен в образцах китайцев хань, но с сильно различающейся частотой:
- 15,0% (6/40) хань из Гуанчжоу (Hammer et al. др. 2005 и Карафет и др. 2001)
- 6,8% (3/44) Хан из Сиань (Хаммер и др. 2005 и Карафет и др. 2001)
- 6,7% (2/30) Хан из Ланьчжоу (Сюэ и др. 2006)
- 3,6% (3/84) Тайваньский Хан (Хаммер и др. 2005)
- 2,9% (1/34) Хан из Чэнду (Сюэ и др. 2006)
- 2,9% (1/35) Хан из Харбин (Сюэ и др. 2006)
- 2,9% (1/35) Хан из округа Мэйсянь (Сюэ и др. 2006)
- 0% (0/32) Хан из города Инин (Сюэ и др. 2006)
Другие популяции, в которых были обнаружены представители N1 * -LLY22g, включают:
- народ хани (4/34 = 11,8%) (Xue et al. 2006)
- люди Sibe (4/41 = 9,8%) (Xue et al. 2006)
- люди Tujia (2/49 = 4,1%) (Хаммер и др. 2005)
- маньчжурцы (2/52 = 3,8% (Хаммер и др. 2005) до 2/35 = 5,7% (Xue et al. 2006)
- Битые люди (1/28 = 3,6%) (Cai 2011) ошибка harv: нет цели: CITEREFCai2011 (help )
- Uyghurs (2/70 = 2,9% (Сюэ и др. 2006) до 2/67 = 3,0%) (Хаммер и др. 2005)
- тибетцы (3/105 = 2,9% (Хаммер и др. 2005) до 3/35 = 8,6% (Сюэ и др. 2006))
- корейцы (0/106 = 0,0% - 2/25 = 8% (Rootsi et al.2006, Xue et al.2006 и Kim 2007 harvnb error: no target: CITEREFKim2007 (help ))
- Вьетнамский народ (2/70 = 2,9%) (Хаммер и др. 2005)
- японцы (0/70 Токусима - 2/26 = 7,7% Аомори ) (Хаммер и др. 2005)
- маньчжурские эвенки (0/26 = 0,0% (Сюэ и др. 2006) до 1/41 = 2,4% (Хаммер и др. 2005))
- алтайцы (от 0/50 северных до 5/96 = 5,2% южных, или от 0/43 бешпельтир до 5/46 = 10,9% кулада), (Хаммер и др.. 2005) (Харьков 2007) harv error: нет цели: CITEREFKharkov2007 (help )
- Shors (2/23 = 8.7%) (Root Си и др. 2006)
- Хакасский народ (5/181 = 2,8%) (Рутси и др. 2006)
- Тувинцы (5/311 = 1,6%) (Рутси и др. 2006)
- Южный Борнео (1/40 = 2,5%) (Rootsi et al. 2006)
- Лесные ненцы (1/89 = 1,1%) (Rootsi et al. al.2006)
- Якуты (0/215 - 1/121 = 0,8%) (Rootsi et al. 2006)
- Турецкий народ (1/523 = 0,2%) (Rootsi et al. 2006) В Турции общее количество субкладов гаплогруппы N-M231 составляет 4% мужского населения.
- Один человек, принадлежащий либо к N *, либо к N1 *, был обнаружено в выборке из 77 мужчин из Катманду, Непал (1/77 = 1,3% N-M231 (xM128, P43, Tat)) (Gayden 2007). Ошибка: нет цели: CITEREFGayden2007 (help )
N1 (xN1a, N1c) был обнаружен в древних костях цивилизации Ляо :
- Niuheliang (Культура Хуншань, 6500–5000 лет назад) 66,7% (= 4/6)
- (, 5000–4200 лет до н.э.) 100,0% (= 12/12)
- (культура нижних сяцзядов, 4200–3600 лет назад) 60,0% (= 3/5)
N1a ( F1206 / M2013 / S11466)
N1a2 -F1008 / L666 clade и N1a1-M46 / Page70 / Tat, по оценкам, имеют одного общего предка в N1a-F1206 / M2013 / S11466 примерно за 15 900 [95% ДИ 13 900 <->17 900] лет до настоящего времени, или 17 621 [95% CI 14,952 <->20,282] лет до настоящего момента.
N1a1 (M46 / Page70 / Tat, L395 / M2080)
Все M46 в базе данных Yfull - M178, т.е. на четверть моложе отделения от F1139.
Мутации, которые определяют субклад N-M46, - это M46 / Tat и P105. Это наиболее частый субклад N. Вероятно, он возник в популяции Северо-Восточной Азии, потому что самые древние образцы соответствуют этому генетическому профилю. N испытал серию узких мест в Сибири и вторичное расширение в Восточной Европе (Rootsi et al. 2006). Гаплогруппе N-M46 примерно 14000 лет.
В Сибири гаплогруппа N-M46 достигает максимальной частоты примерно 90% среди якутов, тюркского народа, проживающего в основном в Республике Саха (Якутия). Однако N-M46 гораздо реже встречается у многих соседей якутов, таких как эвенков и эвенов. Он также был обнаружен в 5,9% (3/51) выборки Hmong Daw из Лаоса (Cai 2011) harv error: no target: CITEREFCai2011 (help ), 2,4% (2/85) выборки из Сеула, Южная Корея (Katoh 2004), ошибка: нет цели: CITEREFKatodiv class="ht"004 (help ) и в 1,4% (1/70) образца из Токусима, Япония (Hammer et al. 2005).
Гаплогруппа N-M46 имеет низкое разнообразие среди якутов, что свидетельствует о узком месте населения или эффекте основателя (Пакендорф 2002) harv error: нет цели: CITEREFPakendorf2002 (справка ). Это было подтверждено исследованием древней ДНК, которое проследило происхождение мужских якутских линий от небольшой группы всадников из Прибайкалья (Crubézy 2010) harv error: no target: CITEREFCrubézy2010 ( справка ).
N1a1a (M178)
Субклад N-M178 определяется наличием маркеров M178 и P298. N-M178 * имеет более высокую среднюю частоту в Северной Европе, чем в Сибири, составляя примерно 60% среди финнов и примерно 40% среди латышей, литовцев 35% среди эстонцев (Derenko 2007 harvnb error: no target: CITEREFDerenko2007 (help ) и Lappalainen 2008 harvnb error: no target: CITEREFLappalainen2008 (справка )).
Мирослава Деренко и ее коллеги отметили, что в этой гаплогруппе есть два подкластера, оба присутствующие в Сибири и Северной Европе, с разной историей. Тот, который они назвали N3a1, сначала распространился на юг Сибири и распространился на Северную Европу. Между тем, более молодой подкластер, который они назвали N3a2, возник на юге Сибири (вероятно, в районе Байкала) (Деренко 2007) harv error: no target: CITEREFDerenko2007 (help ).
N-M178 также был обнаружен в двух На-Дене говорящих Тлыжчо в Северной Америке.
Неолитические образцы из Байкальского региона дали множество Образцы yDNA N и один образец из Фофоново, Бурятия, 5000–4000 гг. до н.э., являются одними из первых образцов Tat в древней летописи.
N1a2 (F1008 / L666)
N1a2a-M128 и N1a2b-B523 / P43, по оценкам, имеют одного общего предка в N1a2-F1008 / L666 примерно 8600 [95% ДИ 7,500 <->9800] лет до настоящего или 9 314 [95% ДИ 7,419 <->11 264] лет до настоящего.
По крайней мере три из шести протестированных мужских особей из «раннего неолита» (охотники-собиратели, использующие керамику) примерно 7200–6200 лет назад) слой на археологическом памятнике Шаманка у южной оконечности озера Байкал был обнаружен как принадлежащий к N1a2-L666.
N1a2a-M128
| Гаплогруппа N-M128 |
|---|
| Возможное место происхождения | Азия |
|---|
| Предок | N1c2 (F1008 / L666) |
|---|
| Определение мутаций | M128 |
|---|
Этот подклад определяется наличием маркера M128. N-M128 был впервые обнаружен в выборке из Японии (1/23 = 4,3%) и в выборке из Центральной Азии и Сибири (1/184 = 0,5%) в предварительном исследовании Y- Вариация ДНК. Впоследствии он был обнаружен с низкой частотой в некоторых выборках из маньчжур, сибэ, эвенков, корейцев, <365.>Хань китайцы, хуэй, тибетцы, вьетнамцы, народ буйей, казахи, Узбеки, уйгуры, салары, Tu, монголы, бузавцы племя калмыков, хакасов и Коми.
Несколько ханьских китайцев, уолед монгол, Цян и тибетец принадлежали к сестринская ветвь (или ветви) N-M128 в парагруппе N-F1154 *.
N1a2b (P43)
Гаплогруппа N-P43 определяется наличием маркера P43. По оценкам, ему около пяти тысяч лет (TMRCA 4,700 [95% ДИ 3,800 <->5,600] лн или 4,727 [95% ДИ 3,824 <->5,693] лет назад). Он очень часто обнаруживается среди северных самодийских народов и носителей обско-угорских языков, а также с низкой или средней частотой наблюдается среди носителей некоторых других уральских языков. языки, тюркские народы, монголы, тунгусские народы и сибирские юпики.
Наибольшая частота N-P43 наблюдается среди популяций северо-западной Сибири: 92% (35/38) в выборке нганасан, 78% (7/9) в выборке энец, 78% (21 / 27) в выборке ханты, 75% (44/59) в выборке тундровых ненцев, 69% (29/42) в другой выборке ненцев, 60% (15/25) в выборка манси, 57% (64/112) в другой выборке хантов, 54% (27/50) в другой выборке нганасан, 45% (40/89) в выборке лесных ненцев, 38% (18/47) в третьей выборке хантов и 25% (7/28) в четвертой выборке хантов. В Европе типы N-P43 имеют самую высокую частоту - 20% среди волго-уральских популяций. Крайней западной границей распространения N-P43 является Финляндия, где эта гаплогруппа встречается лишь с предельной частотой - 0,4%. Тем не менее, N-P43 довольно часто встречается среди вепсов (17,9%), небольшого финского населения, живущего в непосредственной близости от финнов, карелов и эстонцев.
Гаплогруппа N-P43 образует два отличительных субкластера гаплотипов STR: азиатский и европейский., последние в основном распространены среди финно-угороязычных народов и родственных народов (Рутси и др. 2006).
N1a2b1-B478
TMRCA N-B478 оценивается в 3007 [95% ДИ 2171 <->3970] лет назад. Это одна из наиболее распространенных гаплогрупп Y-ДНК среди коренных народов северо-запада Сибири: 69,0% (29/42) ненцев, 50,0% (25/50) нганасанов, 22,2% (12/54) долганов с Таймыра, 7,0% ( 3/43) селькупское, 1,6% (1/63) обско-угорское. Он также довольно распространен среди популяций Средней Сибири, Южной Сибири, Монголии и Кыргызстана: 17,9% (17/95) тувинцев, 15,5% (27/174) хакасов, 13,0% (6/46) тувинцев тожу, 8,7% (2/23) шорцев, 8,3% (2/24) эвенов, 8,2% (5/61) алтайцев, 5,3% (3/57) эвенков, 5,0% (19/381) монголов, 4,9% (3/61) Сарт-Калмак (Кыргызстан), 4,2% (9/216) Якутский, 2,1% (1/47) Торгут (Монголия), 1,4% (1 / 69) Дербет (Калмыкия), 0,9% (1/111) Бурятский. Территориально удаленный член был обнаружен в выборке из Чувашей (1/114 = 0,88%).
Karafet et al. (2018) наблюдали N-P63, который, по-видимому, примерно филогенетически эквивалентен N-B478, в 91,2% (31/34) нганасан, 63,8% (30/47) тундровых ненцев, 42,7% (35/82) лесных ненцев, 14,0% (8/57) долган, 7,0% (9/129) селькупов, 3,3% (3/91) эвенков, 2,7% (2/75) монголов, 2,6% (2/78) коми, 2,5% (2 / 80) Бурятский, 2,0% (2/98) Алтай-Кижи. Эта гаплогруппа не наблюдалась в выборках юкагирцев (0/10), коряков (0/11), телеутов (0/40), кетов (0/44), якутов (0/62) или хантов (0/165).
N1b (F2930)
Гаплогруппа N1b была преимущественно обнаружена у народа И, тибето-бирманской этнической группы в юго-западном Китае, которая произошла от древних племен Цян на северо-западе Китая. Однако он также был обнаружен у людей по всему Китаю и в некоторых образцах людей из Польши, Беларуси, России, Монголии, Кореи, Японии, Вьетнама, Камбоджи, Бутана и Индии.
N2 (Y6503)
N2 (Y6503 / FGC28528; B482 / FGC28394 / Y6584) - первичная ветвь гаплогруппы N-M231, в настоящее время представлена в основном субкладом, N-FGC28435, это распространилось, вероятно, когда-то в первой половине второго тысячелетия нашей эры и было обнаружено у жителей Сербии, Хорватии, Боснии и Герцеговины, Черногория и Турция (Стамбул ).
N-Y7310 (или N-F14667) включает N-FGC28435 и, вероятно, происходит от общего предка, который некоторое время жил в первая половина прошлого тысячелетия. Однако члены N-Y7310 (xFGC28435) имеют более широкий географический диапазон, включая человека из Ростовской области России и венгерского человека с родословной из Сучава, Буковина.
Другие ветви N-P189 включают членов из Италии, Румынии, Словакии и Англии (Девон ). Самый недавний общий предок всех вышеупомянутых сохранившихся N-P189 родословная восходит к до некоторого времени во втором, третьем или даже во второй половине четвертого тысячелетия до нашей эры. Археологический образец, относящийся к ботайской культуре северного Казахстана четвертого тысячелетия до нашей эры, может принадлежать к ветви, предшествующей N-P189.
Линии, принадлежащие к N-Y6503 (xP189) и только отдаленно связаны (время от последнего общего предка оценивается более чем за 10000 лет до настоящего времени) с вышеупомянутыми членами N-P189 были обнаружены у человека из современной Республики Алтай и, вероятно, также в археологическом образце, относящемся к культуре Мезочшат железного века на территории нынешней Венгрии (примерно 2900 лет назад), и в археологическом образце, приписываемом китойской культуре керамики, использующей собиратели из территория вокруг озера Байкал (примерно 6700 лет до настоящего времени).
Древние народы
Образец, обнаруженный на раскопках на стоянке Хоутаомуга в районе Юнхэ поселка Хонганцзы, Даань, Цзилинь, Китай, датируемый 7430-7320 лет назад (Фаза II ранний неолит) принадлежит к гаплогруппе N Y-ДНК и гаплогруппе B4c1a2 мтДНК . Эта выборка аутосомно идентична популяциям неолитического периода бассейна реки Амур, наиболее близким современным представителем которой являются нивхи. Поскольку в статье была обнаружена эта родословная у образца терминального плейстоцена USR1 на Аляске, предполагается, что из Амура в Америку произошел поток генов у населения, принадлежащего к гипотетической чукотско-камчатско-нивхской лингвистической семье.
N также был найден во многих образцах человеческих останков эпохи неолита, эксгумированных из цивилизации Ляо на северо-востоке Китая и в околобайкальской области на юге Сибири. Таким образом, предполагается, что яДНК N, несущая предков уральских народов и тюрко-якутских народов, возможно, возникла в Прибайкалье 8000–6000 лет назад.
Филогения
Филогенетическое дерево
В следующем дереве номенклатура трех источников разделена косой чертой: Дерево ISOGG 10 декабря 2017 г. (версия 12.317)
- NO-M214
- N-M231 / Page91, M232 / M2188
- N1-Z4762 / CTS11499 / L735 / M2291
- N1a-L729
- N-Z1956
- N-Y149447 Китай (Шэньси )
- N1a1- M46 / Page70 / Tat
- N1a1a-M178
- N1a1a1-F1419
- N1a1a1a-L708
- N1a1a1a1-P298
- N1a1a392a
- N1a1a1a1a1-CTS10760
- N1a1a1a1a1a-CTS2929 / VL29 Часто встречается среди литовцев, латышей, эстонцев, северо-запад русские, шведы саамы, карелы, ненцы, финны и марис, средняя частота среди прочих россиян, Белорусы, украинцы и поляки, низкая частота среди коми, мордва, татар, чуваши, долганы, вепсы, селькупы, караногайцы и башкиры
- N1a1a1a1a1a1 -Z4908
- N-Y46443
- N-Y46443 * Россия (Московская область )
- N-BY33095 Россия (Самарская область )
- N1a1a1a1a1a1a-L550 / S431
- N-L550 * Швеция (Округ Кроноберг )
- N-Y9454 Швеция (Округ Вестра-Гёталанд, Округ Эребру ), Финляндия, Россия
- N-Y20911 Финляндия (провинция Западная Финляндия, провинция Южная Финляндия)
- N-Y7795
- N-Y7795 * Швеция (Графство Норрботтен, Седерманланд Округ )
- N-Y29766 Норвегия (Хедмарк )
- N-Y20918 Швеция (Округ Эстергётланд, Округ Вестерботтен ), Финляндия (Провинция Западная Финляндия)
- N-Y28771
- N-Y61225 Швеция (Графство Сёдерманланд, Округ Упсала )
- N-Y30126
- N-Y30126 * Швеция (Округ Эстергётланд )
- N-Y29764
- N-Y29764 * Швеция (Округ Стокгольм )
- N-Y30123 Швеция (Округ Вестерботтен ), Финляндия
- N-S9378
- N-S9378 * Швеция (Округ Вестра-Гёталанд )
- N-S18447 Швеция (Округ Седерманланд, Округ Упсала )
- N-Y36282
- N-Y36282 * Финляндия (провинция Южная Финляндия), Эстония (Ида-Вирумаа )
- N-BY21957 Польша (Куявско-Поморское воеводство ), Швеция (Округ Стокгольм ), Россия (Псковская область ), Финляндия (Южная Финляндия)
- N-FGC14542
- N-FGC14542 * Финляндия (Восточная Финляндия Провинция)
- N-BY21938 Норвегия (Хедмарк ), Швеция (Графство Даларна )
- N-Y17113 Соединенное Королевство (Саффолк, Шотландия) ), Канада (Манитоба ), Норвегия (Вестфолд, Бускеруд ), Швеция (Округ Вестра-Гёталанд )
- N-Y4341
- N-BY21874 Швеция (Округ Сёдерманланд ), Финляндия
- N-Y4338
- N-Y4338 *
- N-Y4339 Швеция (средневековая Сигтуна )
- N-Y12104
- N-Y12104 * Швеция (Округ Эстергётланд )
- N-Y12103 Финляндия (Западная Финляндия) Провинция), Швеция, Норвегия (Hedmark )
- N-Y19111
- N-Y57577 Швеция (Округ Сконе )
- N-Y22774 Финляндия (провинция Западная Финляндия)
- N- Y5611
- N-Y5611 * Англия
- N-Y21546 Швеция (Округ Стокгольм, Округ Вестманланд )
- N-F1983 Швеция (Округ Евлеборг) ), Россия (Липецкая область )
- N-Y10932
- N-Y85136 Швеция (Уезд Упсала )
- N-Y10931 Россия
- N1a1a1a1a1a1a1-L1025 / B215 Наивысший частота среди литовцев, значительная у латышей и эстонцев и меньшая частота у белорусов, украинцев, юго-запад русские и поляки. За исключением эстонцев, L1025 имеет самую высокую долю среди клад N-M231 в ранее упомянутых популяциях. Также наблюдается в Финляндии и Швеции, единичные случаи в Норвегии, Германии, Нидерландах, Соединенное Королевство, Азорские острова, Чехия и Словакия.
- N-L1025 * Россия (Курская область )
- N-BY30389 Швеция (Округ Вестерноррланд ), Финляндия (Провинция Западная Финляндия)
- N-Y13982
- N-A11940
- N-Y140872 Швеция (Уезд Седерманланд, уезд Норрботтен), Финляндия (Южная Финляндия)
- N-Y143451 Россия (Башкортостан )
- N-Y5580
- N-Y93996 Литва (Шяуляйский уезд ), Польша (Подляское воеводство )
- N-BY158 Россия (Воронежская область, Смоленская область, Татарстан ), Беларусь (Витебская область ), Литва (Вильнюсский уезд, Шяуляйский уезд, Паневежский уезд ), Польша (Подляское воеводство.), Украина (Полтавская область ), Казахстан (Жамбылская область )
- N-Z16975
- N-VL69 Беларусь (Минская область ), Россия ( Брянская область ), Казахстан (Костанайская область )
- N-Z16976
- N-Z16976 * Беларусь (Гродненская область ), Украина (Хмельницкая область )
- N-Y13475 Польша, Литва
- N-Y21578 Литва (Каунасский уезд, Тельшяйский уезд ), Россия (Смоленская область )
- N-Y6129 Польша (Варминско-мазурское воеводство ), Литва
- N-Y19113 Польша (Варминско-мазурское воеводство ), Литва (Таурагский уезд )
- N-Y134492 Беларусь (Гродненская область, Минская область, Могилевская область ), Литва, США (Иллинойс )
- N-L551
- N-L551 * Литва (Вильнюсский уезд )
- N-Y15251 Литва (Алитусский уезд ), Польша (Подляское воеводство )
- N-Y46313 Латвия
- N-Y86578 Россия (Белгородская область )
- N-Y14152
- N-BY21911 Польша, Латвия (Краславский р-н ct ), Украина (Киевская область ), Финляндия (Южная Финляндия)
- N-Y13979 Литва, Россия (Рязанская область ), Германия
- N-Y4706
- N-Y4706 * Швеция (графство Стокгольм, графство Сёдерманланд, графство Вестра-Гёталанд), Германия (Нижняя Саксония ), Финляндия (провинция Южная Финляндия), Россия (Татарстан )
- N-A705 Швеция (Округ Кальмар, Округ Эстергётланд )
- N-BY21893 Польша (Великопольское воеводство )
- N-Y139030 Швеция (Округ Вестерботтен ), Norway (Østfold )
- N-Y183040 Russia (Vologda Oblast, Tatarstan)
- N-Y4707 Finland (Western Finland Province,Åland, Southern Finland Province, Eastern Finland Province, Oulu Province)
- N-Z16981
- N-A2358 England, Finland, Latvia (Daugavpils apriņķis )
- N-CTS8173 Estonia (Lääne-Virumaa )
- N-BY32524 Estonia (Järvamaa ), Finland (Oulu Province)
- N-FGC39882 Lithuania (Telšiai County )
- N-A11470 Russia (Tyumen Oblast ), Lithuania (Panevėžys County ), Finland (Southern Finland Province), Netherlands (South Holland,North Brabant )
- N-Y15922 Finland (Southern Finland Province), Estonia (Ida-Virumaa,Pärnumaa ), Latvia (Valmiera,Liepāja,Talsu apriņķis ), Russia (Pskov Oblast ), Poland (Pomorskie )
- N-Y6075 Poland (Małopolskie,Mazowieckie ), Ukraine (Lviv Oblast ), Slovakia, Czech Republic (Moravian-Silesian Region ), United States (New Jersey )
- N-Y11882
- N-Y11882* Russia (Tatarstan)
- N-ZS11617 Russia (Yaroslavl Oblast ), Lithuania
- N-Y24601 Latvia
- N-Y94659 Lithuania, Belarus (Minsk Region )
- N-Y32725
- N-FT96305 Latvia (Krāslava District ), Estonia (Viljandimaa )
- N-Y33333 Ukraine (Sumy Oblast ), Belarus (Brest Region )
- N1a1a1a1a1a2-CTS9976
- N-L1022
- N-Y19098
- N-Y19098* Sweden (Västernorrland County )
- N-A12258 Finland (Eastern Finland Province)
- N-BY117178 Finland (Eastern Finland Province)
- N-Y5004
- N-Y7300
- N-A17632
- N-Y128024 Sweden (Östergötland County )
- N-A16017
- N-A16017* Estonia (Raplamaa )
- N-A16526
- N-FT216144 Finland (Western Finland Province)
- N-Y95406 Estonia (Tartumaa ), Russia (Leningrad Oblast )
- N-Y7297 Finland (Western Finland Province, Southern Finland Province, Lapland Province)
- N-Y15813
- N-Y15813* Finland (Western Finland Province)
- N-Y49008 Finland (Western Finland Province, Oulu Province)
- N-Y46886 Finland (Western Finland Province)
- N-Y15812
- N-A14187 Finland (Western Finland Province), Estonia (Läänemaa )
- N-Y24617 Finland (Western Finland Province), Sweden (Dalarna County ), Russia (Kursk Oblast )
- N-Y23576
- N-Y23576* Finland (Eastern Finlan d Province)
- N-Y15615
- N-BY100801 Sweden (Dalarna County), Finland (Western Finland Province)
- N-A17838 Finland (Western Finland Province, Southern Finland Province), Sweden (Stockholm County )
- N-Y23179 Finland (Western Finland Province, Oulu Province)
- N-Y5005
- N-Y22106
- N-Y47789 Finland (Western Finland Province)
- N-Y79341 Finland (Western Finland Province), Sweden (Västra Götaland County,Värmland County )
- N-Y10756
- N-A13656 Finland (Western Finland Province, Lapland Province), Sweden (Dalarna County )
- N-PH2196
- N-CTS11122 Finland (Eastern Finland Province)
- N-A17277 Finland (Western Finland Province, Southern Finland Province)
- N-PH547 Finland (Lapland Province, Eastern Finland Province, Southern Finland Province), Sweden (Västerbotten County,Norrbotten County )
- N-Y5003
- N-Y5003* Estonia (Hiiumaa ), Finland (Southern Finland Provinc e), United Kingdom (Westminster )
- N-BY22001 Estonia (Läänemaa ), Finland (Western Finland Province), Sweden (Norrbotten County)
- N-BY6007 Russia (Lipetsk Oblast )
- N-Y132182 Sweden (Värmland County ), Estonia (Viljandimaa )
- N-Y20917 Sweden (Kronoberg County ), Finland (Western Finland Province, Southern Finland Province)
- N-Y24502 United Kingdom (Scottish Borders,Calderdale,Kirklees )
- N-Y18420 Finland (Southern Finland Province, Western Finland Province)
- N-Z35267 Finland (Western Finland Province,Åland Province, Southern Finland Province), Estonia (Pärnumaa )
- N-Y6599 Finland (Eastern Finland Province, Oulu Province, Southern Finland Province), Estonia (Jõgevamaa,Hiiumaa ), Russia (Tula Oblast )
- N-Y24000
- N-Y24001 Швеция (Округ Вестерботтен ), Норвегия (Трумс )
- N-A17082 Финляндия (провинция Западная Финляндия, провинция Южная Финляндия), Эстония
- N-Y16503 Финляндия (провинция Западная Финляндия), Провинция Южная Финляндия, провинция Восточная Финляндия, провинция Оулу), Швеция (Графство Евлеборг )
- N-BY6010 США (Вирджиния )
- N-CTS3451
- N-CTS3451 * Русский
- N-Y3667
- N-BY33087
- N-CTS657
- N-CTS657 * Финляндия, Россия (Республика Карелия, Новгородская область)
- N-BY70437 Финляндия (провинция Восточная Финляндия)
- N-BY6024 Финляндия (провинция Восточная Финляндия)
- N-Y45925 Россия (Республика Карелия)
- N-Y26750
- N-Y26750 * Россия (Архангельская область )
- N-A16653 Финляндия (провинция Восточная Финляндия)
- N-PH3568 Финляндия (Провинция Оулу, Восточный Финл и провинция)
- N1a1a1a1a1b-PH1266 / Y28526 / F4134
- N-Y46746 Россия
- N-Y32732
- N-Y32732 * Россия (Санкт-Петербург )
- N-Y192174
- N-Y32731 Россия (Республика Коми ), Швеция
- N1a1a1a1a1c-B479 Нанай, Ульчи
- N1a1a1a1a2- Z1936, CTS10082 Наиболее часто встречается у финнов, вепсов, карелов, шведов саамов, северо-западных русских, башкиры и татары Поволжья, умеренная встречаемость среди прочих русских, коми, ненец, обь -Угры, долганы и сибирские татары, и низкая частота среди мордва, нганасан, чувашей, эстонцы, латыши, украинцы и караногайцы
- N1a1a1a1a2a1c-PH3340 / Y13850
- N1a1a1a1a2a1c1-L>
- N-Y28538
- N-Y28538 * Россия (Ханция-Мансия )
- N-L1032 Россия (Ханция-Мансия), Казахстан (E аст Казахстанская область )
- N-L1442 Венгрия
- N-FT12605 Россия (Липецкая область ), Греция (Аркадия )
- N-Y23732 Россия (Башкортостан )
- N-Y24222
- N-Y24222 * Россия (Татарстан)
- N-Y62987 Россия (Башкортостан, Самарская область )
- N1a1a1a1a2a1c2-Y24361
- N -Y24361 *
- N-Y24360 Россия (Татарстан)
- N1a1a1a1a2a-Z1934
- N-Y159520
- N-Y18421
- N-Y18421 * Россия (Тамбовская область)
- N-Y19110
- N-Y180247
- N-Y180247 * Россия (Татарстан )
- N-FT52835 Россия (Тульская область, Москва )
- N-Y19108
- N-Y19108 * Россия (Татарстан, Кировская область )
- N-Y19453
- N-Y19453 * Россия (Татарстан)
- N-Y19451 Финляндия (провинция Западная Финляндия), Россия (Республика Карелия, Тверская область )
- N-Z1928 / CTS2733
- N-YP6091
- N-YP6 091 * Россия (Иркутская область )
- N-YP6094
- N-Y129131
- N-Y129131 * Россия (Костромская область )
- N-A25107 Россия (Кировская область, HGDP Русский )
- N-YP6092
- N-YP6092 * Россия (Вологодская область )
- N-B195
- N1a1a1a1a2a-Z1925
- N-Z1925 * Швеция ( Округ Норрботтен )
- N-Y29767 Швеция (Округ Норботтен )
- N-Y62904 Финляндия (Провинция Западная Финляндия, Провинция Оулу )
- N1a1a1a1a2a2a1a1-Z1926
- N-Y20920
- N-Z1927
- N-Y22108 Финляндия (провинция Восточная Финляндия, провинция Южная Финляндия, провинция Западная Финляндия), Швеция (Графство Стокгольм, Лен Эребру )
- N-Z1933
- N-CTS4329
- N-Y22091
- N-Y22091 * Норвегия (Нурланд ), Финляндия (Провинция Лапландия )
- N-Y47623
- N-Y47623 * Финляндия (Лапландия)
- N-Y106419 Норвегия (Финнмарк )
- N-Y31247
- N-Y31247 * Россия (Тверская область )
- N-Y31249
- N-Y90283 Финляндия (провинция Лапландия)
- N-Y31244 Финляндия (провинция Западная Финляндия, провинция Восточная Финляндия )
- N-YP6269
- N-YP6269 * Финляндия (провинция Западная Финляндия)
- N-Y151660
- N-Y151660 * Финляндия (провинция Восточная Финляндия)
- N-BY149208 Швеция (Округ Норрботтен )
- N-CTS8565
- N-BY18114 Финляндия (провинция Восточная Финляндия)
- N-BY30394 Финляндия (провинция Восточная Финляндия), Россия (Удмуртская Республика )
- N-Y30513
- N-Y30513 * Финляндия (провинция Южная Финляндия )
- N-Y29759
- N-Y29759 * Финляндия (провинция Лапландия)
- N -Y29758 Швеция (округ Норрботтен)
- N-Z4998
- N-Z4998 * Финляндия (провинция Южная Финляндия)
- N-FGC65190 Финляндия (провинция Южная Финляндия, провинция Восточная Финляндия)
- N-Y18414 Финляндия (провинция Западная Финляндия)
- N-Y20910 Финляндия (провинция Западная Финляндия)
- N-BY194138 Финляндия (провинция Южная Финляндия)
- N-Y28547 Финляндия (провинция Восточная Финляндия)
- N-BY22141 Финляндия (провинция Западная Финляндия, провинция Восточная Финляндия), США (Вашингтон)
- N-FT5834 Финляндия (провинция Западная Финляндия, провинция Южная Финляндия)
- N-BY190112 Финляндия (провинция Западная Финляндия, провинция Южная Финляндия)
- N-Y19097 Финляндия ( Провинция Восточная Финляндия, провинция Южная Финляндия, провинция Западная Финляндия, провинция Оулу)
- N-CTS3223 Финляндия (провинция Южная Финляндия, провинция Восточная Финляндия, провинция Западная Финляндия, провинция Оулу, провинция Лапландия), Швеция (округ Норрботтен, Лен Вестерботтен, Лен Даларна, Лен Вермланд, Лен Сконе), Россия (Ленинградская область )
- N-VL62 Россия (Алтайский край, Кост Ромская область ), Эстония (Харьюмаа ), Финляндия
- N-VL62 * Россия (Челябинская область ), Финляндия
- N- VL60
- N-VL72
- N-Z1939
- N-Y132561
- N-Y132565 Финляндия (провинция Восточная Финляндия)
- N-Y152012
- N-Y152012 * Финляндия
- N- Y152010 Россия (Ленинградская область)
- N-Z2445
- N-Z35031 Финляндия (провинция Восточная Финляндия, провинция Западная Финляндия), Швеция (уезд Вестра-Гёталанд, уезд Вестманланд)
- N-BY28931
- N-Y125841 Финляндия (провинция Восточная Финляндия, провинция Западная Финляндия)
- N-Y11631 Финляндия (провинция Западная Финляндия, провинция Восточная Финляндия, провинция Южная Финляндия, провинция Оулу), Россия (Ленинградская область), Швеция (округ Норрботтен)
- N-CTS7189
- N-BY142665 Финляндия (провинция Восточная Финляндия)
- N-Y37149 Финляндия (провинция Восточная Финляндия)
- N -Y23568 Финляндия (провинция Восточная Финляндия)
- N-Y80226 Финляндия (провинция Восточная Финляндия)
- N-FT20730 Финляндия (провинция Восточная Финляндия), Россия (Республика Карелия)
- N-BY62666 Финляндия (провинция Восточная Финляндия), Норвегия (Осло), США (Вирджиния, Арканзас)
- N-Y24218
- N-Y24218 * Финляндия (провинция Лапландия, провинция Оулу, Провинция Восточная Финляндия)
- N-Y24217 Финляндия (провинция Западная Финляндия, провинция Восточная Финляндия), Россия (Башкортостан )
- N-BY22038 Финляндия (провинция Восточная Финляндия, провинция Южная Финляндия, провинция Западная Финляндия)
- N-Y13974 Финляндия (провинция Южная Финляндия, провинция Восточная Финляндия, провинция Оулу, провинция Западная Финляндия)
- N-Y135981 Финляндия (провинция Восточная Финляндия, провинция Оулу, провинция Южная Финляндия, Западная Финляндия) Prov ince), Норвегия (Финнмарк )
- N-Z4747
- N-Y17790 Финляндия (провинция Восточная Финляндия, провинция Южная Финляндия, провинция Западная Финляндия)
- N-Z1941
- N-BY22090 Финляндия (провинция Южная Финляндия, провинция Восточная Финляндия)
- N-Y21575 Финляндия (провинция Западная Финляндия, провинция Южная Финляндия, провинция Восточная Финляндия, провинция Оулу, провинция Лапландия), Россия (Ленинградская область, Москва, Татарстан )
- N-Z1940 Финляндия (провинция Западная Финляндия, провинция Южная Финляндия, провинция Восточная Финляндия, провинция Оулу, провинция Лапландия), Россия (Ленинградская область), Швеция (уезд Вермланд)
- N1a1a1a1a3-B197 / Y16323
- N1a1a1a1a3a-F4205 Часто встречается среди бурят и цаатанов, умеренно встречается среди Караногайцы, тувинцы, тоджинцы и монголы, и низкая частота среди алтайцев, сибирских татар, казахи, эвенки, крымские татары, каракалпаки, узбеки и украинцы
- N-F4205*
- N-Y16221
- N-Y16221 * Китай (Внутренняя Монголия )
- N-Y16220
- N-Y16220 * Россия (Бурятская Республика )
- N-Y65550 Узбекистан, Турция
- N-F22331 Турция (Мерсин ), Польша (Малопольское воеводство )
- N-Y16312
- N-Y16312 * Узбекистан
- N-Y16320 Казахстан
- N1a1a1a1a3b-B202 Найдено с высоким частота среди чукчей, коряков и сибирских эскимосов
- N1a1a1a1b-M2118
- N1a1a1a1b-M2118 * Эстония
- N-M2016
- N1a1a1a1b1-M1982 / M1993 Якутия (Якут, Эвен, Эвенк )
- N-M1932 Якутия (Якут, Эвен )
- N-M1991
- N-M1988 Якутия (Якут )
- N-Y25011 Якутия (Якут, эвенки )
- N1a1a1a1b2-A9408 Ливан
- N-Y70200 Кореец, Китай
- N-PH1612
- N1a1a1a2-B211 Удмурт, Коми, чуваши, обско-угры, мари, мордва, алтайцы, белорусы, Караногай, карель, башкир, татарин, русский, хакас
- N1a1a1a2a- B181
- N-Y182299 Россия (Кировская область и др.)
- N-Y23788 Россия (Башкортостан, Республика Коми и т. д.)
- N1a1a1a2b-Y23181
- N-Y24317
- N-Y24317 * (xB187) Индия (Андхра мусульманин)
- N1a1a1b -B187 хакасы, шорцы, алтайцы, тувинцы, тожу тувинцы, татары, Башкир
- N1a1a2-Y23747 Япония, Даур, Китай, Тибет (Шигадзе )
- N1a1a2a-Y23749 Япония
- N1a1a2b-Y125664
- N1a1a2b1-MF38607 Аньхой, Хунань
- N1a1a2b2-F22150
- N-F22150 * Хэбэй <709574>>Орокен
- N-MF15288 Хэйлунцзян, корейский
- N-MF55680 Корейский, шигадзе (тибетский )
- N-F1360
- N-F1360 * Китай (Шэньси )
- N1a2-F1008 / L666
- N-F1101
- N-F1101 * Китай (Шаньдун )
- N-F1154
- N-F1154 * Китай (Ляонин )
- N-Y23741
- N-Y23741 * Китай
- N1a2a-M128
- N-M128 * Япония (Фукуи )
- N-Y23738 Вьетнам
- N-CTS1350
- N-CTS1350 * Китай (Ляонин, Тяньцзинь)
- N-F1998
- N-F1998 * Китай (Пекин, Хан, Хэнань), Япония (Кумамото )
- N-MF1633 Китай (Гуандун, Чжэцзян)
- N-MF41795 Корея (провинция Северная Чолла )
- N-Y71703
- N-Y71703 * Япония (Сайтама )
- N-Y173843
- N-Y173843 * Южная Корея
- N-Y174089 Китай (Шаньдун)
- N1a2b-B523 (P43)
- N1a2b-Y126204, VL67 / Z35079, BY29083
- N-VL67 / Z35079
- N1a2b1-B478 (P63) ненцы, нганасаны, долганы, тувинцы, тоджинцы, хакасы, шорцы, эвены, алтайцы, селькупы, эвенки, монголы (сарт-калмак, торгут, дербет, буряты), якуты, обь -Угры, чуваши
- N1a2b1a-B168 эвены
- N1a2b1b-B169
- N1a2b3-B525 Турция, татары, башкиры, казахи, монголы (синьцзян-калмык, монгольский Торгут), Словакия, Болгария, украинцы, белорусы, русские
- N-B525 * Афганистан, араб
- N-BY173686 Россия (Воронежская область )
- N-VL83 Турция
- N-Y136502 Россия (Татарстан ), Болгария ( Добрич )
- N-Y37153
- N-Y37153 * Россия (Татарстан )
- N-BY29131
- N-BY29131 * Турция (Конья ), Словакия (Жилинский район )
- N-BY30476 Россия (Татарстан, Пензенская область )
- N1a2b2-FGC10872 / Y3195
- N1a2b2a- FGC10847 / Y3185 (L1419) Вепсас, Марис, Русские (Архангельская область ), Комис, Пермский край, Республика Коми, обско-угры, чуваши, татары, башкиры, карелы, провинция Западная Финляндия, тувинцы, буряты, хакасы, нганасаны, Азиатские эскимосы
- N1a2b2b-Y23786
- N1b-F2905
- N1b1-CTS582
- N1b1a-Y6374 / Z8029
- N1b1a1-CTS7324 Пекин
- N1b1a2-L727
- N1b1b-Y23789 / CTS4309 Ирак
- N1b1b * -Y23789 Пекин (Хан), Фуцзянь
- N1b1b1-Y60861 Гуанси
- N1b2-M1819 / N-M1897 / CTS12473 / F1173 Китай, Российская Федерация
- N-M1897 * Сычуань
- N-M1928
- N-M1928 * Сычуань (Хан)
- N-Y125475
- N-Y63516 Китай, Корея (провинция Северная Хамгён )
- N-Y193396 Китай, Сингапур
- N-CTS4714
- N-CTS4714 * Наси
- N-F2407
- N-Y24191
- N-Y24191 * Шаньдун
- N-Y24193
- N-Y24193 * Хэбэй
- N-Y24190 Китай, Япония
- N-M1877
- N-M1877 * Шигадзе (тибетский)
- N-F1486 Чунцин
- N-F1486 * Соединенное Королевство (Телугу ), Монголия
- N-M1812
- N-F1260
- N-F1260 * Hubei (Han)
- N-Y62067
- N-Y62067 * Гуандун
- N-Y62132 Гонконг, Китай
- N-M1823
- N-M1823 * Чунцин
- N-M1811
- N-M1811 * Пекин (Хан), Гуанси
- N-Y24355
- N-Y24355 * Китай, Макао, Хошимин (Kinh)
- N-Y64234 Гуандун
- N2-Y6503
- N2-Y6503 * Республика Алтай
- N2a-P189.2
- N2a1-Y6516
- N2a1-Y6516*
- N2a1a-Y7310
- N2a1a-Y7310 * Румыния (венгерский из Сучава )
- N2a1a1-Y7313
- N2a1a1-Y7313*
- N2a1a1a-BY35494 Россия (Ростовская область )
- N2a1a1b-FGC28435
- N2aG1a1 Стамбул), Сербия, Черногория (Плужине), Босния и Герцеговина (Республика Сербская ), Хорватия
- N2a1a1b1-FGC28483 Сербия
- N2a2-Y101945 Соединенное Королевство (Девон ), Россия (Московская область )
Филогенетическая история
Pri или до 2002 г. в академической литературе было по крайней мере семь систем именования Филогенетического дерева Y-хромосомы. Это привело к значительной путанице. В 2002 году основные исследовательские группы объединились и сформировали Консорциум Y-хромосомы (YCC). Они опубликовали совместный документ, в котором было создано единое новое дерево, которое все согласились использовать. Позже группа гражданских ученых, интересующихся популяционной генетикой и генетической генеалогией, сформировала рабочую группу для создания любительского дерева, стремясь быть, прежде всего, своевременным. В приведенной ниже таблице собраны все эти работы на основе знакового Дерева YCC 2002 года. Это позволяет исследователю, просматривающему ранее опубликованную литературу, быстро перемещаться между номенклатурами.
| YCC 2002/2008 (Сокращение) | (α) | (β) | (γ) | (δ) | (ε) | (ζ) | ( η) | YCC 2002 (от руки) | YCC 2005 (от руки) | YCC 2008 (от руки) | YCC 2010r (от руки) | ISOGG 2006 | ISOGG 2007 | ISOGG 2008 | ISOGG 2009 | ISOGG 2010 | ISOGG 2011 | ISOGG 2012 |
|---|
| 12 | VIII | 1U | 25 | Eu16 | H5 | F | N* | N | N1 | N1 | - | - | - | - | - | - | - |
| 12 | VIII | 1U | 25 | Eu16 | H5 | F | N1 | N1 | N1a | N1a | - | - | - | - | - | - | - |
| 12 | VIII | 1U | 25 | Eu16 | H5 | F | N2 | N2a | N1b1 | N1b1 | - | - | - | - | - | - | - |
| 12 | VIII | 1I | 26 | Eu13 | H5 | F | N3* | N3 | N1c | N1c | - | - | - | - | - | - | - |
| 16 | VIII | 1I | 26 | Eu14 | H5 | F | N3a* | M178 | N1c1 | N1c1 | - | - | - | - | - | - | - |
| 16 | VIII | 1I | 26 | Eu14 | H5 | F | N3a1 | N3a1 | N1c1a | N1c1a | - | - | - | - | - | - | - |
Источники Следующие исследовательские группы в соответствии с их публикациями были представлены в создании Дерева YCC.
- αJobling Tyler-Smith 2000 ошибка harvnb: нет цели: CITEREFJoblingTyler-Smitdiv class="ht"000 (help ) и Kaladjieva 2001 ошибка harvnb: нет цели: CITEREFKaladjieva2001 (help )
- βUnderhill 2000 ошибка harvnb: нет цели: CITEREFUnderhill2000 (help )
- γHammer 2001 ошибка harvnb: нет цели: CITEREFHammer2001 (help )
- δKarafet et al. 2001
- εSemino 2000 ошибка harvnb: нет цели: CITEREFSemino2000 (help )
- ζSu 1999 ошибка harvnb: нет цели: CITEREFSu1999 (help )
- ηCapelli 2001 ошибка harvnb: нет цели: CITEREFCapelli2001 (help )
Ненадежные мутации (SNP и UEP)
Делеция b2 / b3 в области AZFc Y-хромосомы, по-видимому, произошла независимо по крайней мере в четырех разных случаях. Следовательно, эту делецию не следует рассматривать как уникальный полиморфизм события, определяющий эту ветвь древа Y-хромосомы (ISOGG 2012).
Генетика
Субклады Y-ДНК N
Древо позвоночника Y-ДНК
Ссылки
Сноски
Цитированная работа
Журналы
- Цай, Сяоюнь; Цинь, Чжендун; Вен, Бо; Сюй, Шухуа; Ван, Йи; Лу, Ян; Вэй, Ланьхай; Ван, Чуанчао; и другие. (2011). О'Рурк, Деннис (ред.). «Миграция людей через узкие места из Юго-Восточной Азии в Восточную Азию во время последнего ледникового максимума, обнаруженного Y-хромосомами». PLOS ONE. 6 (8): e24282. Bibcode : 2011PLoSO... 624282C. doi : 10.1371 / journal.pone.0024282. PMC 3164178. PMID 21904623.
- Кьярони, Жак; Андерхилл, Питер А.; Кавалли-Сфорца, Лука Л. (2009). «Разнообразие Y-хромосомы, человеческое развитие, дрейф и культурная эволюция». Труды Национальной академии наук. 106 (48): 20174–79. Bibcode : 2009PNAS..10620174C. DOI : 10.1073 / pnas.0910803106. PMC 2787129. PMID 19920170.
- Крубези, Эрик; Амори, Сильвен; Кейзер, Кристина; Буаказе, Кэролайн; Боднер, Мартин; Гиберт, Морган; Рёк, Александр; Парсон, Вальтер; Алексеев Анатолий; Лудес, Бертран (2010). «Эволюция человека в Сибири: от замороженных тел к древней ДНК». BMC Evolutionary Biology. 10 : 25. doi : 10.1186 / 1471-2148-10-25. PMC 2829035. ПМИД 20100333.
- Деренко Мирослава; Малярчук, Борис; Денисова, Галина; Возняк, Марцин; Гжибовски, Томаш; Дамбуева Ирина; Захаров, Илья (2007). «Распространение гаплогруппы N Y-хромосомы из Южной Сибири в Европу». Журнал генетики человека. 52 (9): 763–70. DOI : 10.1007 / s10038-007-0179-5. PMID 17703276.
- Гайден, Тензин; Кадены, Алисия М.; Регейро, Мария; Сингх, Нанда Б.; Животовский, Лев А.; Андерхилл, Питер А.; Кавалли-Сфорца, Луиджи Л.; Эррера, Рене Дж. (2007). «Гималаи как направленный барьер для потока генов». Американский журнал генетики человека. 80 (5): 884–94. DOI : 10.1086 / 516757. PMC 1852741. PMID 17436243.
- Хаммер, Майкл Ф.; Карафет, Татьяна М.; Парк, Хваён; Омото, Кейчи; Харихара, Синдзи; Стоункинг, Марк; Хораи, Сатоши (2005). «Двойное происхождение японцев: общая почва для Y-хромосом охотников-собирателей и фермеров». Журнал генетики человека. 51 (1): 47–58. DOI : 10.1007 / s10038-005-0322-0. PMID 16328082.
- Илумяэ (2016). «Гаплогруппа N хромосомы Y человека: нетривиальная разрешенная во времени филогеография, которая пересекает языковые семьи». Американский журнал генетики человека. 99 (1): 163–73. doi : 10.1016 / j.ajhg.2016.05.025. PMC 5005449. PMID 27392075.
- Кан Ху (2015). «Дихотомическая структура гаплогруппы N». arXiv : 1504.06463 [q-bio.PE ].
- Карафет, Татьяна; Сюй, Липин; Ду, Руофу; Ван, Уильям; Фэн, Ши; Wells, R.S.; Редд, Алан Дж.; Зегура, Стивен Л.; Хаммер, Майкл Ф. (2001). «История отцовского населения Восточной Азии: источники, закономерности и микроэволюционные процессы». Американский журнал генетики человека. 69 (3): 615–28. DOI : 10.1086 / 323299. PMC 1235490. PMID 11481588.В этой статье образец "Южный Хань" исследовательской группы Карафета и Хаммера описывается как происходящий из провинции Гуандун, а образец "Северная Хань" описывается как происходящий из Шэньси.
- Карафет, ТМ; Mendez, F. L.; Meilerman, M. B.; Андерхилл, П. А.; Zegura, S.L.; Хаммер, М. Ф. (2008). «Новые бинарные полиморфизмы изменяют форму и увеличивают разрешение дерева гаплогруппы Y-хромосомы человека». Геномные исследования. 18 (5): 830–38. doi : 10.1101 / gr.7172008. PMC 2336805. PMID 18385274.
- Karafet, T. M.; Проба, B.; Cox, M. P.; Sudoyo, H.; Дауни, S.; Lansing, J. S.; Хаммер, М. Ф. (2010). «Основное деление Восток-Запад лежит в основе стратификации Y-хромосомы в Индонезии». Молекулярная биология и эволюция. 27 (8): 1833–44. doi : 10.1093 / molbev / msq063. PMID 20207712.
- Като, Тору; Мунхбат, Батмунх; Тунаи, Кеничи; Мано, Шухей; Андо, Харуэ; Оюнгерел, Ганджуур; Чаэ, Гуэ-Тэ; Хан, Хуун; Цзя, Гуань-Цзюнь; Токунага, Кацуши; Мунхтувшин, Намид; Тамия, Ген; Иноко, Хидетоши (2005). «Генетические особенности монгольских этносов, выявленные с помощью анализа Y-хромосомы». Ген. 346 : 63–70. doi : 10.1016 / j.gene.2004.10.023. ПМИД 15716011.
- Харьков, В.Н.; Степанов, В. А.; Медведева, О. Ф.; Спиридонова, М.Г.; Воевода, М. И.; Тадинова, В. Н.; Пузырев, В. П. (2007). «Различия генофонда между северными и южными алтайцами, выведенные из данных о гаплогруппах Y-хромосомы». Российский журнал генетики. 43 (5): 551–62. DOI : 10.1134 / S1022795407050110. PMID 17633562. S2CID 566825.
- Ким, Ук; Ю, Таг-Гын; Ким, Сон Чжу; Шин, Донг-Джик; Тайлер-Смит, Крис; Джин, Хан-Джун; Квак, Кён-Дон; Ким, Ын-Так; Пэ, Юн-Сон (2007). Благосклонный Михаил (ред.). «Отсутствие ассоциации между Y-хромосомными гаплогруппами и раком простаты у корейского населения». PLOS ONE. 2 (1): e172. Bibcode : 2007PLoSO... 2..172K. doi : 10.1371 / journal.pone.0000172. PMC 1766463. PMID 17245448.
- Lappalainen, T.; Laitinen, V.; Salmela, E.; Andersen, P.; Huoponen, K.; Savontaus, M.-L.; Лахермо, П. (2008). «Миграционные волны в регион Балтийского моря». Анналы генетики человека. 72 (3): 337–48. DOI : 10.1111 / j.1469-1809.2007.00429.x. PMID 18294359. S2CID 32079904.
- Малярчук, Борис; Деренко, Мирослава; Гжибовски, Томаш; Лункина, Арина; Чарны, Якуб; Рычков, Серж; Морозова Ирина; Денисова, Галина; Miscicka-Sliwka, Danuta (2004). «Дифференциация митохондриальной ДНК и Y-хромосом в российских популяциях». Человеческая биология. 76 (6): 877–900. doi : 10.1353 / hub.2005.0021. PMID 15974299. S2CID 17385503.
- Пакендорф, Бриджит; Морар, Бхарти; Тарская, Лариса; Кайзер, Манфред; Судьял, Химла; Родевальд, Александр; Стоункинг, Марк (2002). «Y-хромосомные доказательства сильного сокращения численности мужского населения якутов». Генетика человека. 110 (2): 198–200. DOI : 10.1007 / s00439-001-0664-4. PMID 11935328. S2CID 30671159.
- Рутси, Сиири; Животовский, Лев А; Балдович, Мариан; Кайзер, Манфред; Кутуев, Ильдус А; Хусаинова, Рита; Бермишева, Марина А; Губина, Марина; и другие. (2006). «Северный путь против часовой стрелки гаплогруппы N Y-хромосомы из Юго-Восточной Азии в Европу». Европейский журнал генетики человека. 15 (2): 204–11. doi : 10.1038 / sj.ejhg.5201748. PMID 17149388.
- Сирак, Кендра; Фернандес, Даниэль; Чероне, Оливия; Харни, Иадаоин; Ма, Мэтью; Маллик, Свапан; Роланд, Надин; Адамски, Николь; Брумандхошбахт, Насрин; Каллан, Кимберли; Кандилио, Франческа; Лоусон, Энн Мари; Мандл, Кирстен; Оппенгеймер, Йонас; Стюардсон, Кристин; Залзала, Фатьма; Андерс, Александра; Бартик, Юрай; Коппа, Альфредо; Дашцевег, Тюмень; Эвингер, Шандор; Фаркаш, Зденек; Хайду, Тамаш; Баярсайхан, Джамсранджав; Макинтайр, Лорен; Моисеев, Вячеслав; Окумура, Мерседес; Пап, Ильдико; Пьетрусевский, Михаил; Рачки, Пал; Шефчакова, Алена; Софикару, Андрей; Szeniczey, Tamás; Сёке, Бела Миклош; Ван Гервен, Деннис; Васильев, Сергей; Белл, Линн; Райх, Дэвид; Пинхаси, Рон (2020). "Слуховые косточки человека как альтернативный оптимальный источник древней ДНК". Геномные исследования. Март (3): 427–436. doi : 10.1101 / gr.260141.119. PMID 32098773.
- Вэнь, Бо; Се, Сюаньхуа; Гао, Сун; Ли, Хуэй; Ши, Хун; Песня, Сюфэн; Цянь, Тинчжи; Сяо, Чуньцзе; и другие. (2004b). «Анализ генетической структуры тибето-бирманских популяций выявляет предвзятую половую принадлежность южных тибето-бирманцев». Американский журнал генетики человека. 74 (5): 856–65. DOI : 10.1086 / 386292. PMC 1181980. PMID 15042512.
- Xue, Y.; Зерджал, Т; Бао, Вт; Чжу, S; Шу, Q; Сюй, Дж; Ду, Р; Fu, S; и другие. (2005). «Демография мужчин в Восточной Азии: контраст между севером и югом во временах роста человеческой популяции». Генетика. 172 (4): 2431–39. doi : 10.1534 / genetics.105.054270. PMC 1456369. PMID 16489223.
Веб-сайты
- ISOGG (2012). «Дерево гаплогруппы Y-ДНК 2012».
- Макдональд, Дуг (2004). «Макдональд Y Гаплогруппы мира» (PDF). Архивировано из оригинального (PDF) 28 июля 2004 г. Проверено 29 августа 2009 г.
Дополнительная литература
Филогенетика
Внешние ссылки
 | Викискладе есть материалы, связанные с гаплогруппой N Y-ДНК. |


 Предполагаемые доисторические маршруты миграции для линии гаплогруппы N Y-хромосомы.
Предполагаемые доисторические маршруты миграции для линии гаплогруппы N Y-хромосомы.  Прогнозируемое распределение субгаплогрупп гаплогруппы N. (A) N * -M231, (B) N1 * -LLY22g, (C) N1a-M128, (D) N1b-P43, (E) N1c-M46.
Прогнозируемое распределение субгаплогрупп гаплогруппы N. (A) N * -M231, (B) N1 * -LLY22g, (C) N1a-M128, (D) N1b-P43, (E) N1c-M46.