
Войти
| Грегаринасина | |
|---|---|
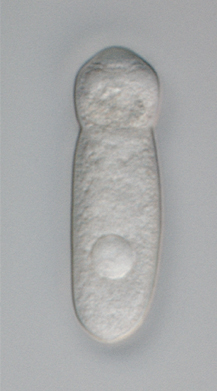 | |
| Живой образец септата (или) грегарина, демонстрирующий характерный «головной» участок трофозоита, содержащий эпимерит на его переднем конце. Септатные грегарины - кишечные паразиты членистоногих. | |
| Научная классификация | |
| Домен: | Eukaryota |
| (без рейтинга): | SAR |
| (без рейтинга): | Альвеолата |
| Тип: | Apicomplexa |
| Класс: | Conoidasida |
| Подкласс: | Gregarinasina |
| Орден | |
| 178>Грегариния | |
грегарины представляют собой группу апикомплексан альвеолатов, классифицируемых как грегарина или грегариния . Крупные (примерно полмиллиметра) паразиты обитают в кишечнике многих беспозвоночных. Они не встречаются ни у каких позвоночных. Однако грегарины тесно связаны как с Toxoplasma, так и Plasmodium, которые вызывают токсоплазмоз и малярию соответственно. Оба протиста используют белковые комплексы, подобные тем, которые образуются грегаринами для скользящей подвижности и проникновения в клетки-мишени. Это делает их отличными моделями для изучения подвижности скольжения с целью разработки вариантов лечения токсоплазмоза и малярии. Ожидается, что у насекомых будут обнаружены тысячи различных видов грегаринов, и 99% этих грегаринов все еще нуждаются в описании. Каждое насекомое может быть хозяином нескольких видов. Одна из наиболее изученных грегаринов - Gregarina garnhami. В целом грегарины считаются очень успешными паразитами, поскольку их хозяева распространены по всей планете.
Грегарины встречаются как в водной, так и в наземной среде. Хотя они обычно передаются по орофекальному маршруту, некоторые передаются с гаметами хоста во время копуляции (Monocystis ).
У всех видов четыре или более спорозоитов (точное число зависит от вида), снабженных апикальным комплексом, ускользают из ооцист, процесс, называемый эксцистацией, находят свой путь в соответствующую полость тела и проникают в клетки-хозяева в их непосредственном окружении. Спорозоиты появляются внутри клетки-хозяина, начинают питаться и развиваются в более крупные трофозоиты. У некоторых видов спорозоиты и трофозоиты способны к бесполому воспроизведению - процесс, называемый шизогония или мерогония. Однако у большинства видов в жизненном цикле отсутствует шизогония.
У всех видов два зрелых трофозоита в конечном итоге объединяются в пары в процессе, известном как сизигия, и развиваются. Во время сизигии ориентация гамонтов различается у разных видов (из стороны в сторону, голова к хвосту). Стена образуется вокруг каждой пары гамонтов, которые затем начинают делиться на сотни гамет. Зиготы образуются в результате слияния двух гамет, которые, в свою очередь, окружаются стенкой ооцисты. Внутри ооцисты происходит мейоз с образованием спорозоитов. Сотни ооцист накапливаются в каждой гаметоцисте, и они выделяются через фекалии хозяина или через смерть и разложение хозяина.
 является кишечным паразитом асцидий. Эти асептные грегарины не имеют эпимеритов и вместо этого обладают органеллами прикрепления, известными как мукроны.
является кишечным паразитом асцидий. Эти асептные грегарины не имеют эпимеритов и вместо этого обладают органеллами прикрепления, известными как мукроны. Сообщается, что грегарины заражают более 3000 видов беспозвоночных.
Грегарины были признаны таксоном от Грасс в 1953 г. Три порядка, на которые они в настоящее время разделены, были созданы Левином и др. в 1980 году.
В настоящее время в этом таксоне известно около 250 родов и 1650 видов. Они делятся на три порядка в зависимости от среды обитания, диапазона хозяев и морфологии трофозоитов.
Большинство видов имеют моноксенный жизненный цикл с участием одного беспозвоночного хозяина. В жизненном цикле стадия внеклеточного питания известна как трофозоит.
Архигрегарины встречаются только в морских местообитаниях. Они обладают кишечными трофозоитами, сходными по морфологии с инфекционными спорозоитами. Филогенетический анализ показывает, что эта группа парафилетична и требует разделения. Обычно в каждой споре этой группы четыре зоита.
Эугрегарины встречаются в морских, пресноводных и наземных средах обитания. Эти виды обладают крупными трофозоитами, которые по морфологии и поведению существенно отличаются от спорозоитов. Этот таксон содержит большинство известных видов грегарина. Кишечные эугрегарины разделяются на перегородку - подотряд Septatorina - и асептат - подотряд Aseptatorina - в зависимости от того, разделен ли трофозоит поверхностно на поперечная перегородка. Асептные виды - это в основном морские грегарины.
Уроспоридианы - асептные эугрегарины, поражающие целомические пространства морских хозяев. Как правило, у них отсутствуют структуры прикрепления и они образуют гамоновые пары, которые свободно пульсируют в целомической жидкости.
. Моноцистиды представляют собой асептные эугрегарины, которые инфицируют репродуктивные пузырьки наземных кольчатых червей. Эти последние виды имеют тенденцию тесно разветвляться с неогрегаринами и, возможно, нуждаются в переклассификации. Обычно в каждой споре этой группы содержится восемь зоитов.
Неогрегарины встречаются только у наземных хозяев. Эти виды имеют пониженное содержание трофозоитов и имеют тенденцию инфицировать ткани, отличные от кишечника. Обычно в каждой споре этой группы по восемь зоитов.
Эугрегарины и неогрегарины различаются по ряду аспектов. Неогрегарины в целом более патогенны для хозяев. Эугрегарины размножаются посредством спорогонии и гаметогонии, в то время как неогрегарины имеют дополнительную шизогенную стадию - мерогонию - в пределах их хозяев. Мерогония может быть внутриклеточной или внеклеточной в зависимости от вида.
Исследования ДНК предполагают, что архигрегарины являются предками других.
Кавалье-Смит предложил значительную переработку этого таксона, предполагая, что полифилий из эвгрегаринов. Он разделил грегарины на три класса. Первый из них - Gregarinomorphea - включает, Cryptosporidiidae и, кроме того, Rhytidocystidae, ранее считавшиеся дивергентными кокцидиями или Apicomplexa incertae sedis. Ортогрегариния с двумя новыми отрядами была создана для грегаринов, наиболее близких к Cryptosporidium. Второй класс - Paragregarea - был создан для архигрегаринов, а новый порядок, который сам был создан для суперсемейства. п. и Велоксидиум. Был создан третий класс - для Filipodium и Platyproteum. Таким образом, эвгрегарины оказались разделенными и распределенными между этими тремя классами вместе с некоторыми другими апикомплексами.
. Эта точка зрения была оспорена в 2017 году Симдьяновым и соавторами, которые провели глобальную комплексный анализ имеющихся морфологических и молекулярно-филогенетических данных и заключение, что эвгрегарины скорее являются монофилетическим таксоном.
Некоторые роды грегаринов в настоящее время не классифицированы:, Грегарина, Нематопсис и.
Паразиты представляют собой относительно крупные веретенообразные клетки по сравнению с другими апикомплексами и эукариотами в целом (некоторые виды имеют размер>850 мкм в длину). У большинства грегаринов есть продольные эпицитарные складки (пучки микротрубочек под поверхностью клетки с нематодоподобным поведением изгиба): вместо этого кренуляции обнаруживаются у уроспоридий.
Грегарины способны перемещаться и изменять направление по поверхности за счет скользящей подвижности без использования ресничек, жгутиков или ламеллиподии. Это достигается за счет использования комплекса актина и миозина. Комплексы нуждаются в актиновом цитоскелете для выполнения их скользящих движений. В предложенной модели «укупорки» нехарактерный белковый комплекс движется назад, продвигая паразитов вперед.
Грегарины - одни из самых старых известных паразитов, описанные врачом Франческо Реди в 1684 году.
Первое формальное описание было сделано Дюфуром в 1828 году. Он создал род Gregarina и описал Gregarina ovata от. Он считал их паразитическими червями. Коелликер признал их простейшими в 1848 году.
| Wikisource содержит текст статьи Британской энциклопедии 1911 года Грегарины. |