
Войти
Генетика и археогенетика Южной Азии - это изучение генетики и археогенетика этносов Южной Азии. Его цель - раскрыть генетическую историю этих групп. Географическое положение Южной Азии делает ее биоразнообразие важным для изучения раннего расселения анатомически современного человека в Азии.
Исследования, основанные на вариациях митохондриальной ДНК (мтДНК ), полученные о генетическом единстве различных различных субпопуляций Южной Азии. Выводы исследований, основанные на вариациях Y-хромосомы и вариабельности аутосомной ДНК, были разными, хотя многие исследователи утверждают, что большинство предковых узлов утверждают, что большинство предковых узлов филогенетического древа всех типов мтДНК возникло в Южной Азии. Недавние исследования генома, по-видимому, показывают, что у жителей Азии являются потомками двух основных предковых компонентов, один из которых ограничен Южной Азией (южно-индийские предки, происходящие от людей IVC, и коренное население Южной Азии, отдаленно связанное с андаманцев ) и другой компонент (предки северных индейцев), происходящие от людей IVC и степняков, что делает его более близким к тем, что находится в Центральной Азии, Азии и Европа. Исследование 2016 года выявило еще два наследственных компонента на материковой части Индии, обозначенные как AAA и ATB, которые являются вызывающими для племен, говорящими на австро-азиатском языке и говорящих на тибето-бирманском языке, соответственно. В исследовании также делается вывод, что популяции архипелага Андаманских островов образуют отдельную родословную, которая «оказалась коанцестральной по отношению к океаническим популяциям». Геномные исследования генетического ландшафта Южной Азии как совокупность представителей населения Южной Азии и Восточной Азии. Компонент восточноазиатского происхождения, обнаруженный в Индии, в основном ограниченными определенными популяциями в предгорьях Гималаев и на северо-востоке Индии.
Было обнаружено, что предковый узел филогенетического дерева всех мтДНК типов (гаплогруппы митохондриальной ДНК ), обычно встречающихся в Центральной Азии, Азии и Европе, а также в Южной Азии с относительно высокими частотами. Предполагаемое расхождение этого общего предкового узла произошло чуть менее 50 000 лет назад. В своих материнских линиях существуют различные субклады M, за которые следуют R и U субклады. На сегодняшний день время слияния этих митохондриальных гаплогрупп приближается к 50 000 лет назад.
Основные отцовские линии, представленные Y-хромосомами, - это гаплогруппы R1a1, R2, H, L и гаплогруппа J2.. Некоторые исследователи утверждали, что гаплогруппа Y-ДНК R1a1 (M17) имеет автохтонное южноазиатское происхождение. Однако предложения о центральноазиатском происхождении R1a1 также довольно распространены.
Все линии мтДНК и Y-хромосомы за пределами Африки соответствуют от трех основателей:
Все эти шесть гаплогрупп-основателей можно найти в современных популяциях Южной Азии. Более того, гаплогруппа M мтДНК и гаплогруппы C и D Y-хромосомы ограничены районом к востоку от Южной Азии. Все популяции западных евразийских происходят из гаплогрупп N и R мтДНК и гаплогруппы F Y-хромосомы.
Эндикотт и др. заявляет, что эти факты согласны с гипотезой единственного исхода из Восточной Африки 65 000 лет назад по южному региону, при этом западно-евразийские линии отделяются от южноазиатских линий где-то между Восточной / Северо-Восточной Африкой и Южной Южной Азия.
Все преобладающие маркеры генома выходцев из Южной Азии западноевразийцами и, возможно, происходят из страны Азии или Южной Азии сама
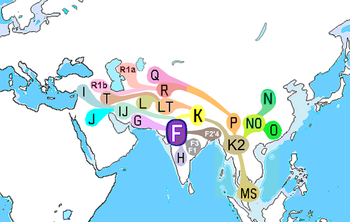
 Гипотетическая карта перемещения человека в ную Азию, основанная на митохондриальной ДНК и источников путях распространения.
Гипотетическая карта перемещения человека в ную Азию, основанная на митохондриальной ДНК и источников путях распространения. Наиболее частые гаплогруппы мтДНК в Южной Азии - это M, R и U (где U является потомком R).
Аргументы в пользу более долгосрочной «конкурирующей модели Y-хромосомы» Стивен Оппенгеймер считает, что Индия является поставщиком евразийских гаплогрупп мтДНК, которые он называет «евразийскими канунами». По мнению Оппенгеймера, весьма вероятно, что почти все человеческие материнские линии в Центральной Азии, на Ближнем Востоке и в Европе произошли только от четырех линий мтДНК, которые возникли в Южной Азии 50 000–100 000 лет назад.
макрогаплогруппа M, которая рассматривается как кластер протоазиатских материнских линий, составляет более 60% южноазиатской мтДНК.
Макрогаплотип M в Индии включает в себя множество подгрупп, которые отличаются от других подлиний в Восточной Азии, особенно монголоидных популяций. Глубокие корни филогении M указывает на указание на реликвию южноазиатских линий по сравнению с другими подсистемами M (в Восточной Азии и в других местах), предполагаемое происхождение этих подгаплогрупп in-situ в Южной Азии, скорее всего, в Индии. Эти глубоко укоренившиеся линии не зависят от языка и распространяются на все языковые группы в Индии.
Практически все современные среднеазиатские линии передачи MtDNA M, похоже, принадлежат к восточно-евразийской (монгольской ), а не южноазиатские подтипы гаплогруппы M, что указывает на то, что крупномасштабная миграция из нынешних тюркских говорящих народов Центральной Азии в Индию не происходила. Отсутствие гаплогруппы M в европейских странах по сравнению с ее столь же высокими показателями среди выходцев Южной Азии, Восточной Азии и некоторых популяций Центральной Азии, контрастирует с западно-евразийскими склонами южноазиатских отцовских линий.
. Предполагается, что сформировано первое время Южного заселения Евразии анатомически современными людьми.
| Гаплогруппа | Важные подклассы | Популяции |
|---|---|---|
| M2 | M2a, M2b | По всему континенту с низким уровнем присутствия на северо-западе. Пик в Бангладеш, Андхра-Прадеш, прибрежном Тамил Наду и Шри-Ланке |
| M3 | M3a | Концентрированный на Северо-Запад Индии. Самый высокий среди парсов Мумбаи |
| M4 | M4a | Пики в Пакистане, Кашмире и Андхра-Прадеше |
| M6 | M6a, M6b | Кашмир и у берегов Бенгальского залива, Шри-Ланка |
| M18 | По всей Азии. Пик в Раджастане и Андхра-Прадеше | |
| M25 | Умеренно часто в Керале и Махараштре, но довольно нечасто в других местах Индии |


 Пространственное распределение M, R и U гаплогрупп мтДНК и их подгаплогрупп в Южной Азии. 362>макрогаплогруппа R (очень большое и старое подразделение макрогаплогруппы N ) также широко представлена и составляет остальные 40% южноазиатской мтДНК. Очень старым и важным ее разделом является гаплогруппа U, хотя и присутствует в регистрации Евразии, несколько субкладов, характерных для Южной Азии.
Пространственное распределение M, R и U гаплогрупп мтДНК и их подгаплогрупп в Южной Азии. 362>макрогаплогруппа R (очень большое и старое подразделение макрогаплогруппы N ) также широко представлена и составляет остальные 40% южноазиатской мтДНК. Очень старым и важным ее разделом является гаплогруппа U, хотя и присутствует в регистрации Евразии, несколько субкладов, характерных для Южной Азии.
Наиболее важные южноазиатские гаплогруппы в R:
| Гаплогруппа | Популяции |
|---|---|
| R2 | Широко распространены по субконтиненту |
| R5 | Широко распространен по большей части Индии.. Пики в прибрежном юго-западе Индии |
| R6 | широко распространены по всей Индии с низкими темпами.. Пики среди тамилов и кашмирцев |
| W | Найдены в северо-западных штатах.. Пики в Гуджарате, Пенджабе и Кашмире, низкая частота в других местах. |
Гаплогруппа U является субгаплогруппой макрогаплогруппы R. из Южной Азии. Южноазиатские линии U наиболее отличаются от европейских, и их слияние с общим предком также насчитывает около 50 000 лет.
| Гаплогруппа | Популяции |
|---|---|
| U2 * | (a парагаплогруппа ) редко распространена, особенно в северной части Южной Азии. Он также находится на юго-западе Аравии. |
| U2a | показывает относительно высокую плотность в Пакистане на северо-западе Индии, но также и в Карнатаке, где он достигает более высокой плотности. |
| U2b | имеет наибольшую концентрацию в Уттар-Прадеше, но также встречается во многих других местах, особенно в Керале и Шри-Ланке. Также встречается в Омане. |
| U2c | особенно важен в Бангладеш и важен Бенгалии. |
| U2l | , возможно, является важным в количественном отношении субкладом в Южной Азии, достигая высокой концентрации (более 10%) в Уттар-Прадеше, Шри-Ланке, Синде и некоторых частях Карнатаки. Это также имеет значение в Омане. гаплогруппа мтДНК U2i названа «западноевразийской» в Бамшад и др. исследования, но «Восточно-Евразийский регион (в основном специфический для Индии)» в Kivisild et al. исследование. |
| U7 | эта гаплогруппа широко представлена в Гуджарате, Пенджабе и Пакистане. Возможная родина этой гаплогруппы охватывает Гуджарат (самая высокая частота, 12%) и Иран, потому что эта частота резко снижается как на восток, так и на запад. |
 Отклонение гаплогруппы F и ее потомков.
Отклонение гаплогруппы F и ее потомков. Основными гаплогруппами ДНК Y-хромосомы Южной Азии являются H, J2, L, R1a1 и R2. Согласно последним исследованиям, их географическое происхождение перечислено следующим образом:
| Основные южноазиатские линии Y-хромосомы: | H | J2 | L | R1a | R2 |
|---|---|---|---|---|---|
| Basu et al. (2003) | без комментариев | без комментариев | без комментариев | Центральная Азия | без комментариев |
| Кивисилд и др. (2003) | Индия | Западная Азия | Индия | Южная и Западная Азия | Южно-Центральная Азия |
| Cordaux et al. al. al. (2004) | Индия | Западная или Центральная Азия | Ближний Восток | Центральная Азия | Юго-Центральная Азия |
| Сенгупта и другие. (2006) | Индия | Ближний Восток и Центральная Азия | Южная Индия | Северная Индия | Северная Индия |
| Тансим и другие. (2006) | Индия | Левант | Ближний Восток | Южная и Центральная Азия | Южная и Центральная Азия |
| Sahoo et al. al. (2006) | Южная Азия | Ближний Восток | Южная Азия | Южная или Западная Азия | Южная Азия |
| Мирабал и другие. (2009) | без комментариев | без комментариев | без комментариев | Северо-Западная Индия или Средняя Азия | без комментариев |
| Чжао и другие. (2009) | Индия | Ближний Восток | Ближний Восток | Центральная Азия или Западная Евразия | Центральная Азия или Западная Евразия |
| Шарма и др. (2009) | без комментариев | без комментариев | без комментариев | Южная Азия | без комментариев |
| Thangaraj et al. (2010) | Южная Азия | Ближний Восток | Ближний Восток | Южная Азия | Южная Азия |
Гаплогруппа H (Y-ДНК) часто встречается в Южной Азии. Сегодня H редко встречается за пределами Южной Азии, распространен романистов, особенно среди подгруппы среди H-M82. H также довольно распространен в древних образцах Европы и до сих пор встречается редко у европейцев и арабов Леванта. Гаплогруппа H часто встречается среди населения Индии, Шри-Ланки, Непала, Пакистана и Мальдив. Все три ветви гаплогруппы H (Y-ДНК) находятся в Южной Азии.
Это ветвь гаплогруппы F и происходит от семьи GHIJK. Гаплогруппа H, как полагают, возникла в Южной Азии между 30 000 и 40 000 лет назад. Вероятное место интродукции - Южная Азия, так как она сосредоточена там. Похоже, что это основная гаплогруппа Y-хромосомы палеолитических жителей Южной Азии. Некоторые особи в Южной Азии также обладают более редким субкладу H3 (Z5857). Гаплогруппа H никоим образом не ограничивается определенными группами населения. Например, H принадлежит примерно 28,8% индоарийских каст. и у племен около 25–35%.
Гаплогруппа J2 присутствовала в Южной Азии в основном как J2a-M410 и J2b-M102 со времен неолита (9500 YBP). Клады J2 достигают пиковых частот в Северо-Западной Индии и находятся на 19% в южноиндийских кастах, 11% в североиндийских кастах и 12% в Пакистане. В Южной Индии присутствие J2 выше среди средних средних (21%), за ним следуют высшие касты (18,6%) и низшие касты (14%). Среди кастовых групп самая высокая частота J2-M172 наблюдается у тамилов веллаларов из Южной Индии - 38,7%. J2 также присутствует в племенах и имеет частоту 11% у австро-азиатских племен. Среди австро-азиатских племен преобладающее значение J2 встречается в Лодхе (35%). J2 также присутствует в южноиндийском уровне горном племени тода на 38,46%, в племени андх из Телангана на 35,19% и в племя кол из Уттар-Прадеш с соотношением 33,34%. Было обнаружено, что гаплогруппа J-P209 более распространена у мусульман-шиитов Индии, из 28,7% принадлежат к гаплогруппе J, с 13,7% в J-M410, 10,6% в J- M267 и 4,4% в J2b (Eaaswarkhanth 2009) harv error: нет цели: CITEREFEaaswarkhanth2009 (help ).
В Пакистане самые высокие частоты J2-M172 наблюдались среди парсов - 38,89%, дравидийцев говорящих брахуи на 28,18% и макрани белуджи на 24%. Он также встречается в 18,18% в Макрани Сиддис и в 3% в Карнатака Сиддис.
J2-M172 из встречается с общей точностью 10,3% среди сингальцев Шри-Ланки. На Мальдивах 20,6% мальдивского населения оказались положительными по гаплогруппе J2.
По словам доктора Спенсера Уэллса, L -M20 возник в регионе Памирский узел в Таджикистане и мигрировал в Пакистан и Индию ок. 30 000 лет назад. Однако в большинстве других исследований предполагалось западноазиатское происхождение L-M20 и связывали его распространение в долине Инда (~ 7000 лет назад) с неолитическими фермерами. Существуют три подветви гаплогруппы L: L1-M76 (L1a1), L2-M317 (L1b) и L3-M357 (L1a2), обнаруженные на разных уровнях в Южной Азии.
Гаплогруппа L показывает время неолитической экспансии. Клада присутствует в индейском населении с общим ок. 7–15%. Гаплогруппа L чаще встречается среди южноиндийских каст (примерно 17–19%) и встречается 68% в некоторых кастах Карнатаки, но несколько реже в касты северной Индии (около 5–6%). Присутствие гаплогруппы L довольно редко среди племенных групп (около 5,6–7%), однако умеренное, 14,6%, присутствующих среди ченчу.
Среди региональных социальных групп, от умеренного до высокого. наблюдались у конканастха-браминов (18,6%), пенджабцев (12,1%), гуджаратцев (10,4%), ламбадитов (17,1%), джатов ( 36,8%)
В Пакистане субклады L1-M76 и L3-M357 L-M20 достигают общих частот 5,1% и 6,8% соответственно. Гаплогруппа L3 (M357) часто встречается среди бурушо (примерно 12%) и пуштунов (примерно 7%). На самой большой частоте встречается в юго-западной провинции Белуджистан вдоль побережья Макрана (28%) дельты реки Инд. L3a (PK3) встречается примерно в 23% нуристанцев на северо-западе Пакистана.
Клада присутствует в умеренном распределении среди населения Пакистана в целом (примерно 14%).
В одном исследовании было обнаружено, что 16% сингальцев имеют положительную реакцию на гаплогруппу L-M20. В другом исследовании было обнаружено, что 18% принадлежат к L1.
В Южной Азии R1a1 часто встречается в ряде демографических групп, а также в самых высоких STR разнообразии, которое заставляет рассматривать его как локус происхождения.
Хотя R1a возник ок. 22 000–25 000 лет назад его субклад M417 (R1a1a1) диверсифицировался ок. 5800 лет назад. Распределение M417-субкладов R1-Z282 (включая R1-Z280) в Центральной и Восточной Европе и R1-Z93 в Азии предполагает, что R1a1a диверсифицировалась в пределах евразийских степей или Ближнего Востока и Кавказ регион. Место происхождения этих субкладов играет роль в дебатах о происхождении индоевропейцев.
В Индии высокий процент этой гаплогруппы обнаружен в брамины регистрации Бенгалии (72%) на восток, Гуджарат Лоханас (60%) на запад, Хатри (67%) на севере, Айенгарские брамины (31%) на юге. Он также был обнаружен у нескольких южноиндийских дравидийских говорящих племен, включая кота (41%) Тамилнада Ченчу (26 %) и Валмики из Андхра-Прадеш, а также Ядав и Каллар из Тамил Наду, предполагая, что M17 широко распространен у этих южных индейцев. племена. Помимо этого, исследования показывают высокий процент в группах, различающихся по регионам, таких как Manipuris (50%) на крайнем северо-востоке и среди пенджабцев (47%) на крайнем северо-западе.
В Пакистане он встречается в 71% среди племен Моханна в провинции Синд к югу и в 46% среди балтитов из Гилгит-Балтистан к северу.
23% сингальцев из 87 субъектов выборки оказались R1a1a (R-SRY1532) положительный согласно исследованию 2003 года.
На Мальдивах было обнаружено, что 23,8% мальдивцев быть R1a1a (M17) положительным.
Люди в Тераи регионе, Непал показывают R1a1a на уровне 69%.
В Южной Азии частота линий R2 и R2a составляет около 10–15% в Индии и Шри-Ланке и 7 –8% в Пакистане. По крайней мере, 90% индивидов R-M124 находятся в Южной Азии. Он также встречается в Кавказ и Средняя Азия с меньшей частотой. Генетическое исследование Mondal et al. 2017 пришел к выводу, что гаплогруппа Гаплогруппа R2 возникла в северной Индии и уже существовала до миграции в степи.
Среди региональных групп она встречается среди Запада. бенгальцы (23%), нью-дели индусы (20%), пенджабцы (5%) и Гуджаратцы (3%). Среди племенных групп кармалицы Западной Бенгалии показали самый высокий уровень (100%), за ними следуют лодхи (43%) на востоке, а бхил в Гуджарат на западесоставляли 18%, Тарас на севере показал это на 17%, Ченчу и Паллан на юге были на 20% и 14% соответственно. Среди кастовых групп высокий процент показывают Джаунпур кшатрии (87%), камма (73%), Бихар Ядав (50%), Хандаят (46%) и Каллар (44%).
Это также значительно выше у многих браминов. группы, включая пенджабских браминов (25%), бенгальских браминов (22%), конканастху браминов (20%), чатурведи (32%), бхаргавы (32%), кашмирские пандиты (14%) и лингаяты брамины (30%).
Мусульмане Северной Индии имеют частоту 19% (суннитов ) и 13% (шиитов ), в то время как мусульмане Давуди Бора в западном штат Гуджарат имеют частоту 16% и мусульмане маппила из Южной Индии имеют частоту 5%.
Гаплогруппа R2 встречается у 14% народа бурушо. Среди народа хунза он встречается у 18%, тогда как парсы показывают его в 20%. Он также обнаружен в северо-восточной части Афганистана.
38% сингальцев Шри-Ланки, согласно исследованию 2003 года, оказались R2-положительными.
12% мальдивцев Мальдив имеют R2.
В Непале R2 проценты представлены от 2% до 26% в разных группах согласно различным исследованиям. Newars демонстрируют широкую распространенность - 26%, в то время как жители Катманду показывают ее на уровне 10%.
Индийский консорциум вариаций генома (2008) делит население Южной Азии на четыре этнолингвистические группы: индоевропейские, дравидийский, тибето-бирманский и австро-азиатский. В исследованиях молекулярной антропологии используются три типа маркеров: вариация митохондриальной ДНК (мтДНК), которая наследуется по материнской линии и является высокополиморфной, вариация Y-хромосомы, которая включает однопародительскую передачу по мужским линиям и вариацию аутосомной ДНК.
Большинство исследований, основанных на вариативности мтДНК, сообщают о генетическом единстве южноазиатских популяций по языку, кастам и племенным группам. Вполне вероятно, что гаплогруппа M была принесена в Азию из Восточной Африки южным маршрутом самой ранней волной миграции 78000 лет назад.
Согласно Kivisild et al. (1999): «Незначительные совпадения с линиями, описанными в других евразийских популяциях, демонстрируют, что недавние иммиграции оказали очень небольшое влияние на врожденную материнскую генофонда выходцев из Южной Азии. Несмотря на различия, наблюдаемые в Индии, эти изменения происходят от ограниченного числа родословных основателей. Эти линии, вероятнее всего, были занесены в Южную Азию в период среднего палеолита, до заселения Европы 48 000 лет назад и, возможно, Старого Света в целом ». Basu et al. (2003) также подчеркивает лежащее в основе единство женских линий в Индии.
Выводы, основанные на вариации Y-хромосомы, были более разнообразными, чем заключение, основанные на вариации мтДНК. Хотя Кивисилд и др. (2003) ошибка harvtxt: нет цели: CITEREFKivisild_et_al.2003 (help ) предлагает древнее и общее генетическое наследие мужских линий в Южной Азии, Bamshad et al. (2001) предполагает сродство между южноазиатскими мужскими линиями и западными евразийцами, пропорциональное высшему кастовому рангу, и помещает высшие касты южных штатов Индии ближе к восточноевропейцам.
Basu et al. (2003) пришли к выводу, что австро-азиатские племенные народы вошли в Индию сначала из Северо-Западного коридора, гораздо позже некоторые из них - через Северо-Восточный коридор. Принимая во внимание, что Kumar et al. (2007) проанализировали 25 южноазиатских австро-азиатских племен и представили сильную отцовскую генетическую связь между сублингвистическими группами австро-азиатского населения Южной Азии. Mukherjee et al. (2001) помещает пакистанцев и северных индейцев между западноазиатскими и центральноазиатскими популяциями, тогда как Cordaux et al. (2004) утверждает, что индийские кастовые популяции ближе к центральноазиатским. Sahoo et al. (2006) и Sengupta et al. (2006) предполагают, что кастовое население Индии не подвергалось каким-либо примесям в последнее время. Сангхамитра Саху завершает свое исследование следующими словами:
Нет необходимости, синхронизировать текущих данных, искать истоки отцовского наследия индийцев во времена оседлого земледелия за пределами Южной Азии. Многолетняя концепция людей, языка и сельского хозяйства, прибывающих в Индию вместе через северо-западный коридор, не выдерживает тщательного изучения. Недавние утверждения о связи гаплогрупп J2, L, R1a и R2 с одновременным происхождением среди представителей различных линий индийских каст из-за пределов Южной Азии отвергают, хотя наши результаты подтверждают местное происхождение гаплогрупп F * и H. Из остальных только J2 указывает на недавний недвусмысленный внешний вклад. из Азии, а не из Центральной Азии. Текущее распределение частот гаплогрупп, за исключением родословных, в основном географических, а не культурными детерминантами. По иронии судьбы, именно на северо-востоке Индии, среди групп туберкулеза, есть четкие доказательства крупномасштабной демической диффузии, прослеживаемой по генам, культуре и языку, но, очевидно, не по сельскому хозяйству.
Анализ ближайшего соседа проведен Mondal et al. 2017 пришел к выводу, что индийские Y-линии близки к южным европейским популяциям и время расхождения между двумя предшествующими степными миграциями. ":
Эти результаты предполагают, что европейское происхождение в индийских популяциях могло быть намного старше и сложнее, чем предполагалось, и может появиться у агрономов первой волны или даже раньше
— Mondal et al., 2017Результаты исследований, основанных на вариабельности аутосомной ДНК, также различались. В крупном исследовании (2009 г.) с использованием более 500 000 двуаллельных аутосомных маркеров Райх выдвинул гипотезу о том Эти две «реконструированные» древние популяции назвал «предками южных индейцев» (ASI) и «предками северных индейцев» (ANI), что современное население Южной Азии возникло в результате смешения двух генетически различных предковых популяций, датируемых постголоценовая эпоха. «Происхождение ANI значительно выше у индоевропейцев, чем у дравидов. Акерс, предполагая, что предки ASI, возможно, говорили на дравидийском языке до, как смешались с ANI». и близок к ближневосточникам, выходцам из Центральной Азии и Европы, ASI не имеет тесного родства с пределами субконтинента. Древняя система ДНК "ASI" недоступна, коренные андаманцы Onge используются в качестве несовершенного заместителя ASI (согласно Райху и др., Андаманцев, хотя и отличаются от них, являются ближайшим живым населением к АСИ). Согласно Райху и др., Предки ANI и ASI встречаются по всему субконтиненту (как на севере, так и на юге Индии) в различных пропорциях, и что «предки ANI колеблются от 39 до 71% в Индии и выше в традиционных верхних регионах. касты и индоевропейские спикеры. «
Мурджани и др. 2013 заявляют, что ASI, хотя и не связан с каким-либо группой,« связаны (отдаленно) с коренными жителями Андаманских островов ». Мурджани и др. Также предполагают возможный приток генов В исследовании сделан вывод о том, что «почти все группы, говорящие на индоевропейских или дравидийских языках, лежат на градиенте различной степени родства с западноевразийцами в ППТС (именой« индейской клиникой »)».
Исследование 2013 года с использованием единого -нуклеотидный полиморфизм (SNP) показывает, что геном андаманцев (онге) ближе к геномам других мировых океанических негрито, чем к геномам жителей Южной Азии.
Согласно Басу и др.., 2016, подробный анализ показал, что геномная структура населения материковой Индии лучше всего объясняется вкладом четырех предковых компонентов. В дополнение к ANI и ASI, Басу и др. (2016) идентифицировали два предковых компонента в материковой Индии, которые явл яются языками для австро-азиатских языков. племен и тибето-бирманских спикеров, которые они обозначили как AAA (от «Предки австро-азиатских») и ATB (от «Предков тибето-бирманцев») соответственно. Исследование также предполагает, что население Андаманских островов архипелаг образуют особую родословную, который «оказался коанцестральным по отношению к океаническим популяциям».
Граница смешения между линиями ANI и ASI датируется периодом c. 4,2–1,9 тыс. Лет назад, Moorjani et al. (2013), что соответствует индийскому бронзовому веку и связывается авторами с процессом деурбанизации цивилизации долины Инда и перемещением населения в систему Ганга в зарождающемся индийском железном веке. Basu et al. (2003) предполагает, что «дравидийские племена, возможно, были широко распространены по всей Индии до прихода кочевников, говорящих на индоевропейском языке» и что «формирование популяций путем деления, которое привело к эффекту на основе и дрейфа, наложило свой отпечаток на генетические структуры современного языка» человека. населения ". Генетик П.П. Маджумдер (2010) утверждал, что результаты Райха и др. (2009) находятся в замечательном соответствии с предыдущими исследованиями с использованием мтДНК и Y-ДНК:
Предполагается, что население Центральной Азии внесло основной с использованием данных о вариациях митохондриальной ДНК, собранные из различных исследований, мы показали, что популяции Центральной Азии и Азии, в генофонд Индии, особенно в генофонд Индии, а также мигранты предположительно переехали в Индию через территорию нынешнего Афганистана и Пакистана. Северная Америка, Индия, Северная Африка, Северная Америка, Индия, Индия. с использованием вышеизложенных недавних исследований с использованием более 500 000 биаллельных маркеров обнаружило градиент генетической близости индейских популяций с западными евразийцами с севера на юг. Эта особенность, вероятно, связана с пропорциями предков, происходящих от генофонда Евразии, как показало это исследование, больше у популяций, населяющих северную Индию, чем тех, которые населяют южную Индию.
Чауби и др. В 2015 году был обнаружен характерный восточноазиатский наследственный компонент, в основном ограниченные конкретные популяциями в предгорьях Гималаев и северо-восточной части Индии. Самая высокая частота этого компонента наблюдается среди тибето-бирманских говорящих групп северо-востока, а также обнаруживается на высоком уровне в андаманских популяциях, с существенным присутствием среди говорящих на австроазиатском языке. Обнаружено, что он в значительной степени отсутствует у индоевропейских и дравидийских носителей, за исключением некоторых конкретных этнических групп, живущих в предгорьях Гималаев и в центральной и южной частях Индии. Однако исследователи предположили, что происхождение ханьцев (прокси для жителей Восточной Азии), измеренное в изученных андаманских группах, связано с частичным улавливанием сродства андаманцев с меланезийцами и малайзийскими негритосами, как было в предыдущем исследовании Chaubey et al. предположил «глубокое общее происхождение» между андаманцами, меланезийцами и другими группами негрито (а также выходцами из Южной Азии) и родство негритосов Юго-Восточной Азии и меланезийцев (а также андаманцев) с жителями Восточной Азии.
Lazaridis и другие. (2016) отмечает: «Демографическое влияние степных популяций на Южную Азию было значительным, поскольку Мала, южноиндийская далитов популяция с минимальным количеством предков Северной Индии (ANI) вдоль ' Предполагается, что индийский клин такой родословной имеет ~ 18% степных предков, в то время как калаши в Пакистане имеют ~ 50% родословную, связанную со степью ". В исследовании Lazaridis et al., Проведенном в 2016 году, оценивается (6,5–50,2%) примесь, связанная со степью, у жителей Южной Азии. Lazaridis et al. далее отмечает, что «полезным направлением будущих исследований является более полный отбор образцов древней ДНК у степных популяций, а также у популяций Центральной Азии (к востоку от Ирана и к югу от степи), что может выявить более близкие источники ANI, чем рассмотренные здесь, а также из Южной Азии для непосредственного определения траектории изменения населения в этом районе.
Патак и др. 2018 пришли к выводу, что индоевропейские носители гангского Равнины и дравидийские носители имеют значительную родословную раннего-среднего бронзового века (Step_EMBA), но не имеют происхождения степей среднего-позднего бронзового века (Steppe_MLBA). С другой стороны, популяции «Северо-Западная Индия и Пакистан» (PNWI) показали значительную предков Steppe_MLBA наряду с предками Yamnaya (Steppe_EMBA). Исследование также отметило, что образцы древней Южной Азии имели значительно более высокий уровень Steppe_MLBA, чем Steppe_EMBA (или Yamnaya). Исследование также показало, что Рорс co uld может использоваться в качестве прокси для ANI.
Дэвид Райх в своей книге 2018 года Who We Are and How We Got Here утверждает, что анализ 2016 года показал, что ASI имеет значительную Компонент предков происходит от иранских фермеров (около 25% их предков), а оставшиеся 75% их предков происходят от коренных южноазиатских охотников-собирателей. Он добавляет, что ASI вряд ли были местными охотниками-собирателями Южной Азии, как было установлено ранее, но населением, ответственным за распространение сельского хозяйства по всей Южной Азии. В случае ANI, иранские фермерские предки составляют 50%, а остальные - из степных групп, относящихся к Ямной.
Narasimhan et al. (2018) аналогичным образом пришли к выводу, что ANI и ASI были сформированы во 2-м тысячелетии до нашей эры. Им предшествовала смесь AASI (древних исконных южноиндийских, т.е. охотников-собирателей, имеющих общий корень с андаманским); и иранские земледельцы, прибывшие в Индию ок. 4700–3000 гг. До н.э., и «должен быть достигнут долины Инда к 4-му тысячелетию до нашей эры». Согласно Нарасимхану и др., Это смешанное население, которое, вероятно, происходит из цивилизации долины Инда, «внесло большой вклад как в ANI, так и в ASI», которые сформировались во 2-м тысячелетии до нашей эры. ANI сформирован из смесей «групп, связанных с Периферией Инда» и мигрантов из степи, в то время как ASI был сформирован из «групп, связанных с Периферией Инда», которые двинулись на юг и смешались дальше с местными охотниками-собирателями. Предполагается, что происхождение населения ASI составляет в среднем около 73% от AASI и 27% от фермеров, связанных с Ираном. Нарасимхан и др. обратите внимание, что образцы из периферийной группы Инда всегда представляют собой смесь одних и тех же двух ближайших источников AASI и иранского земледельческого происхождения; «Одним из индивидов с периферии Инда, имеющим ~ 42% -ное происхождение по шкале AASI, а у двух других людей - ~ 14-18% -ное происхождение по шкале AASI» (с остальной частью их предков из иранского населения, связанного с земледелием)). Исследовало, что местные охотники-собиратели AASI предоставили расходующуюся ветвь, которая отделилась в то же время, когда предки аборигенов Восточной Азии отделились друг от друга. Заключил, что за короткий промежуток времени к линиим, ведущим к AASI., Выходцы из Восточной Азии, онге и австралийцы.
Генетическое исследование, проведенное Йельменом и др. (2019), показывает, что современные южноазиатские популяции, как правило, наиболее близки к западноевразийцам. Они пришли к выводу, что современные жители Южной Азии представляют собой смесь коренного южноазиатского генетического компонента и более позднего западно-евразийского компонента (происходящего из западной Азии, так и из западных степей). Авторы также утверждают, что коренной южноазиатский генетический компонент отличается от андаманцев, и что андаманцы являются таким образом, это несовершенный прокси. Этот компонент (когда он представлен андаманским онге) был обнаружен в северном индийском гуджарати, и поэтому южноиндийский племенной пания народ (группа преобоми коренная южноазиатская родословная) бы служить лучшим представителем представителем, чем андаманцы (онге), для «коренного южноазиатского» компонента у жителей Южной Азии.
Два генетических исследования (Shinde et al. 2019 и Нарасимхан и др. 2019), проанализировали останки цивилизации долины Инда (части Северо-Западной Индии и Восточного Пакистана бронзового века), обнаружил, что у них смешанная родословная: Шинде и др. содержат около 50-98% генома народов, связанных ранними иранскими фермерами, и 2-50% их генома от южноазиатских охотников-собирателей, имеющих общую родословную с андаманцами, с иранскими родственниками. родословная в среднем преобладает. Образцы, проанализированные Narasimhan et al. 45–82% имели родословную, связанную с иранскими фермерами, а 11–50% имели родословную AASI (или андаманскую родословную, связанную с охотниками и собирателями). В проанализированных образцах обоих исследований практически отсутствует компонент «Степное происхождение », связанный с более поздними индоевропейскими миграциями в Индию. Авторы представили соответствующее количество этих предков значительно различается у разных людей, и пришли к выводу, что необходимо больше образцов, чтобы получить полную картину истории индейского населения.
Исследования Watkins et al. (2005) и Kivisild et al. (2003) на основе аутосомных маркеров пришли к выводу, что индейские кастовые и племенные популяции общее происхождение. Редди и др. (2005) значительно большее генетическое расстояние между кастами и племенами, что указывает на генетическую изоляцию племен и каст.
Viswanathan et al. (2004) в исследовании генетической структуры и родства между племенными популяциями южной Индии заключает: «Генетическая дифференциация высокая, генетические данные не были коррелированы с географическими расстояниями. Таким образом, генетический дрейф, вероятно, значительная роль в формировании генетической изменчивости. наблюдается в племенных популяциях южных индейцев. В остальном, анализ соотношения популяций проявляется, что все индейские и южноазиатские популяции по-прежнему похожи друг на друга, независимо от типичных характеристик, и не проявляет какого-либо особого сходства с африканцами. Мы заключаем, что фенотипическое сходство некоторых групп индейцев и африканцев не отражают тесную связь между этими группами, но лучше объясняются конвергенцией ».
Исследование 2011 года, опубликованное в Американские журнале генетики человека, указывает на то, что индейские предки являются результатом более сложной демографической истории, чем считалось ранее. По мнению исследователей, Южная Азия имеет два основных наследственных компонента, один из которых является сопоставимой с генетическим разнообразием в Центральной Азии, Азии и Европы; другой компонент более ограничен Южной Азией. Однако, если исключить возможность крупномасштабной индоарийской передачи, эти результаты предполагают, что генетическая близость обоих индейских предковых компонентов является генетическим генов на протяжении тысяч лет.
| Викицитатник содержит цитаты, связанные с: Генетика и археогенетика Внешней Азии |