
Войти
 Графики смешения современных популяций Западной Европы, основанные на семи компонентах:. Юг / Запад Европы, Север / Восток Европы. Ближний Восток Южная Азия Восточная Азия Северная Африка / Африка к югу от Сахары
Графики смешения современных популяций Западной Европы, основанные на семи компонентах:. Юг / Запад Европы, Север / Восток Европы. Ближний Восток Южная Азия Восточная Азия Северная Африка / Африка к югу от Сахары 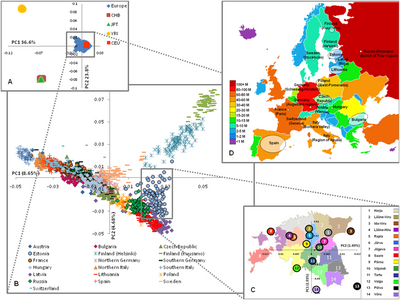 Европейская генетическая структура (на основе 273 464 SNP). Структура три уровня, выявленные ПК-анализом: А) межконтинентальный; Б) внутриконтинентальный; и C) внутри одной страны (Эстония), где показаны медианные значения PC1 и 2. D) Европейская карта, иллюстрирующая происхождение выборки и размер популяции. CEU - штата Юта с происхождением из Северной и Западной Европы, CHB - китайцы хань из Пекина, JPT - японцы из Токио и YRI - йоруба из Ибадана, Нигерия.
Европейская генетическая структура (на основе 273 464 SNP). Структура три уровня, выявленные ПК-анализом: А) межконтинентальный; Б) внутриконтинентальный; и C) внутри одной страны (Эстония), где показаны медианные значения PC1 и 2. D) Европейская карта, иллюстрирующая происхождение выборки и размер популяции. CEU - штата Юта с происхождением из Северной и Западной Европы, CHB - китайцы хань из Пекина, JPT - японцы из Токио и YRI - йоруба из Ибадана, Нигерия. генетическая история из Европы, начиная с верхнего палеолита, неотделима от более широкой Евразии. Примерно 50-40 000 лет назад (50-40 тыс. Лет назад) из недифференцированной «неафриканской » линии возникла базальная западно-евразийская линия (включая с отдельной восточноазиатской линией). 70-50 тыс. И восточные, и западные евразийцы приобрели примесь неандертальцев в Европе и Азии.
Европейские линии раннего современного человека (EEMH) между 40 и 26 тыс. Лет назад (ориньяк ) все еще существовали часть большого западноевразийского «мета-населения», связанного с популяциями Центральной Азии. Дивергенция в генетически обособленных субпопуляциях в пределах страны Евразии является результатом воздействия основанного давления и эффектов основателя во время Последнего ледникового максимума (LGM, Gravettian ). К концу LGM, после 20 тыс. Лет назад, западноевропейская линия, получившая название западноевропейских охотников-собирателей (WHG), возникла из солютриан рефугиума во время Европейский мезолит. Эти мезолитические культуры охотников-собирателей в степени заменены в неолитической революции появлением линий ранних европейских представителей (EEF), происходящих от мезолитического населения Западной Азии (Анатолия <241).>и Кавказ ). В европейском бронзовом веке в некоторых частях Европы снова произошли существенные замены в результате вторжения древних северо-евразийских (ANE) линий из понско-каспийских степей. Эти за ущерб населения бронзового века связаны с культурой кубков археологически и с индоевропейской экспансией лингвистически.
В результате перемещений населения во время мезолита в Бронзовый век, современные европейские популяции отличаются различиями в происхождении WHG, EEF и ANE. Нормы примеси изменяются географически; в позднем неолите происхождение WHG у фермеров в Венгрии составляло около 10%, в Германии - около 25%, а в Иберии - до 50%. Вклад EEF более значителен в Средиземноморской Европе и снижается в направлении северной и северо-восточной Европы, где происхождение WHG сильнее; сардинцы считаются ближайшей европейской группой к населению ВЭФ. Происхождение ANE встречается по всей Европе, с максимумом около 20% у балтийских людей и финнов.
Этногенез современных этнических групп Европы в исторический период связан с многочисленными примесью событий, в первую очередь связанных с римским, германским, норвежским, славян, арабским и турецкий расширения.
Исследования генетической истории Европы стали возможны во второй половине 20 века, но не результаты с высоким разрешением до 1990-х годов. В 1990-х годах стали возможны предварительные результаты, но они остались в основном ограниченными исследованиями линий митохондрий и Y-хромосомы. Аутосомная ДНК стала более доступной в 2000-х, а с середины 2010-х результатов ранее недостижимого разрешения, многие из которых были основаны на полногеномном анализе древней ДНК, опубликованы в ускоренном темпе.
предыстория европейских народов может быть прослежена путем изучения археологических памятников, лингвистических исследований и исследования ДНК людей, живущих в Европе, или древней ДНК. Исследования продолжаются, и занимаются и падают. Хотя можно отследить перемещение людей по Европе с помощью анализа ДНК основателя, большая часть информации об этих перемещениях получена из археологии.
Важно отметить, что заселение Европы не происходило миграциями, поскольку может показаться предложенным. Скорее, процесс заселения был сложным и «вероятным, происходящим происходящим потоком генов».
Из-за естественного отбора процента ДНК неандертальцев в древние европейцы со временем постепенно уменьшались. С 45 000 до 7 000 BP процент упал с 3–6% до 2%. Удаление аллелей, происходящих от неандертальцев, происходило чаще вокруг генов, чем других частей генома.
Неандертальцы еще 130 000 лет назад населяли большую часть Европы и Западной Азии. Они существовали в Европе еще 30 000 лет назад. В конце концов их заменили анатомически современные люди (AMH; иногда известные как кроманьонцы ), которые начали появляться в Европе 40 000 лет назад. Предположим, что два вида гоминидов, вероятно, сосуществовали в Европе, антропологи давно задавались вопросом, использовали ли эти два вида. Вопрос был решен только в 2010 г., когда было установлено, что в евразийских популяциях присутствует примесь неандертальцев, оцениваемая в среднем в 1,5–2,1%. Возник вопрос, имело ли место это смешение в Европе или, скорее, в Леванте до передачи AMH в Европу.
Были также предположения о наследовании определенных генов от неандертальцев. Например, один локус MAPT 17q 21.3, который разделен на глубокие генетические линии H1 и H2. Линия H2, по-видимому, ограничена европейскими популяциями, несколько авторов выступали за наследование от неандертальцев, начиная с 2005 г. Однако предварительные результаты секвенирования полного генома неандертальцев в то время (2009 г.) не показали доказательств межпородного скрещивания между неандертальцами и современными людьми. К 2010 году данные Сванте Паабо (Институт эволюционной антропологии Макса Планка в Лейпциге, Германия), Ричарда Э. Грина (Калифорнийский университет, Санта-Крус) и Дэвида Райха (Гарвардская Медицинская школа), сравнивая генетический материал трех неандертальцев с генетическим опытом пяти современных людей, действительно показала связь между неандертальцами и современными людьми за пределами Африки.
 Замена неандертальцев ранними современными людьми
Замена неандертальцев ранними современными людьми Считается, что современные люди начали населять Европу в период верхнего палеолита около 40 000 лет назад. Некоторые данные свидетельствуют о распространении культуры ориньяк.
С чисто патрилинейной точки зрения Y-хромосома, возможно, что старая гаплогруппа C1a2, F и / или E могут быть самыми старыми в Европе. Они были найдены в некоторых очень старых человеческих останках в Европе. Однако другие гаплогруппы гораздо чаще встречаются среди ныне живущих европейских мужчин.
Гаплогруппа I (M170), которая сейчас относительно широко распространена в Европе, может представлять собой маркер палеолита - ее возраст оценивается примерно в 22000 лет назад. Хотя сейчас он сконцентрирован в Европе, он, вероятно, возник у мужчин с Ближнего Востока или Кавказа или от их ближайших потомков, c. 20–25 000 лет назад, когда он отклонился от своего непосредственного предка, гаплогруппы IJ. Примерно в это же время появилась культура верхнего палеолита, известная как Граветтиан.
. Более ранние исследования Y-ДНК вместо этого были сосредоточены на гаплогруппе R1 (M173): самой многочисленной линии среди ныне живущих. Европейские мужчины; Считается также, что R1 появился ~ 40 000 лет назад в Центральной Азии. Однако теперь считается, что R1 появился значительно позже: исследование 2008 года датировало самого общего общего предка гаплогруппы IJ 38 500, а гаплогруппы R1 18 000 лет назад. Это свидетельствует о том, что колонисты гаплогруппы IJ создали первую волну, а гаплогруппа R1 появилась намного позже.
Таким образом, генетические данные предполагают, что, по крайней мере, с точки зрения отцовской родословной, отдельные группы современных людей пошли в Европу двумя путями: с Ближнего Востока через Балканы и еще один из Центральной Азии через Евразийскую степь, к северу от Черного моря.
Мартин Ричардс и др. представ, что 15-40% использования клонов мтДНК восходят к миграциям палеолита (в зависимости от того, допускает ли один из них несколько событий основателя). Гаплогруппа U5 мтДНК, датируемая ~ 40–50 кЯ, прибыла во время первой ранней колонизации верхнего палеолита. По отдельности он составляет 5–15% от общего числа линий мтДНК. Средний U.P. движения отмечены гаплогруппами HV, I и U4. HV разделился на Pre-V (возраст около 26000 лет) и более крупную ветвь H, обе из которых распространились по Европе, возможно, благодаря контактам Граветта.
Гаплогруппа H составляет около половины генных линий в Европе, причем многие подгруппы. Вышеупомянутые линии мтДНК или их предшественники, скорее всего, прибыль в Европу через Ближний Восток. Это контрастирует с данными ДНК Y , согласно которому около 50% плюс мужских линий характеризуются суперсемейством R1, которое, возможно, имеет центральноазиатское происхождение. Орнелла Семино постулирует, что эти различия могут быть частично связаны с очевидным более поздним молекулярным возрастом Y-хромосом по другим локусами, что предполагает более быструю замену предыдущих Y-хромосом. Гендерные различия в миграционном демографическом поведении также будут влиять на наблюдаемые модели мтДНК и Y-вариация ».
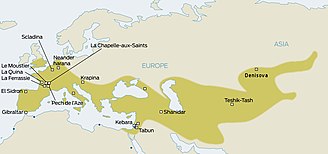
 Европейские убежища LGM, 20 тыс. Лет назад. солютрейские и прото-солютрейские культуры Эпи-граветтианская культура
Европейские убежища LGM, 20 тыс. Лет назад. солютрейские и прото-солютрейские культуры Эпи-граветтианская культура Начало последнего ледникового максимума («LGM») c. 30 тыс. Лет до н. Э., В конце МИС 3, что привело к депопуляции Северной Европы. Согласно классической модели, люди находили убежище в климатических заповедниках (или рефугиумах) следующим образом:
Это событие увеличило общее генетическое разнообразие в Европе, "результат дрейфа, согласующийся с предполагаемым узким местом населения во время последнего ледникового максимума». Когда ледники отступили примерно 16000–13000 лет назад, Европа начала медленно заселяться людьми из рефугиумов, оставляя генетические сигнатуры.
Некоторые представители гаплогруппы Y отошли от своих родительских гаплогрупп где-то во время или вскоре после LGM.Гаплогруппа I2 распространена на западных Балканах, а также в остальной части юго-восточной и центрально-восточной Европы с более умеренными частотами.. Его частота быстро падает в Центральной Европе, что позволяет предположить, что выжившие, несущие линии I2, распространились преимущественно через Юго-Восточную и Центрально-Восточную Европу.
Чинниоглу видит доказательства существования анатолийского убежища, в котором также находилась Hg R1b1b2. Сегодня R1b доминирует в ландшафте y-хромосомы в Европе, включая Британские острова, что указывает на то, что могли быть большие изменения в составе населения, основанные на миграциях после LGM.
Семино, Пассарино и Перичич помещают истоки гаплогруппы R1a в украинское убежище ледникового периода. Его нынешнее распространение в Восточной Европе и некоторых частях отражает повторное заселение Европы из южных российских / украинских степей после максимума позднего оледенения.
С точки зрения мтДНК, Ричардс и др. представится, что большая часть разнообразия мтДНК в Европе объясняется послеледниковыми повторными экспансиями во время позднего верхнего палеолита / мезолита. «Региональный анализ в некоторой степени предположение о том, что большая часть Центральной Европы была заселена в основном из юго-запада, когда климат улучшился. Участвующие линии включают большую часть наиболее распространенной гаплогруппы H, а также большую часть K, T, W и X. "В ходе исследования не удалось определить, были ли в этот период новые линии линий мтДНК с Ближнего Востока; значительный вклад был сочтен маловероятным. 51>
Альтернативная модель большего числа представилась Билтоном и др.
Из исследования 51 человека исследователи идентифицировали пять отдельных генетических кластеров древних Европейцы во время LGM: Вестоницкий кластер (34000–26000 лет назад), связанный с культурой граветтов ; кластер Мальта (24 000–17 000 лет), связанный с культурой Мальта-Бурет, кластер Эль-Мирон ( 19 000–14 000 лет назад), связанное с Магдаленская культура ; скопление Виллабруна (14000–7000 лет назад) и скопление Сацурблия (13000–10000 лет назад)).
Примерно 37000 лет назад все древние европейцы начали свое существование разделить некоторые п. редки с современными европейцами. Эта популяция-основательница представлена GoyetQ116-1, 35000-летним экземпляром из Бельгии. Эта линия снова исчезает из записи и не встречается до 19 000 лет назад в Испании в Эль-Мироне, что свидетельствует о сильном родстве с GoyetQ116-1. В течение этого интервала отчетливое скопление Вестонице преобладает в Европе, даже в Гойе. Повторное расширение скопления Эль-Мирон совпало с потеплением после отступления ледников во время Последнего максимума ледников. От 37000 до 14000 лет назад население Европы состояло из изолированной популяции, происходящей от основной популяции, которая не скрещивалась значительно с другими популяциями.
Мезолит (после LGM) значительно разошлись из-за их относительной изоляции в течение нескольких тысячелетий, из-за жесткого давления отбора во время LGM и из-за эффектов основателя, вызванных быстрым расширением из убежищ LGM в начало мезолита. К концу LGM, примерно от 19 до 11 тыс. Руб. Лет назад, появились знакомые разновидности евразийских фенотипов. Тем не менее, родословная мезолитических охотников-собирателей Европы (WHG) не сохранилась в качестве основного вклада в какое-либо современное население. Скорее всего, они были голубоглазыми и сохранили темную пигментацию кожи, как до LGM EEMH. Варианты HERC2 и OCA2 для голубых глаз продолжаются от линии WHG, также были обнаружены у людей ямная.
Около 14000 лет назад кластер Виллабруна отошел от сродства GoyetQ116-1 и начал проявлять больше сродства с Ближнего Востока, сдвиг, совпал с повышением температуры межстадиального периода Бёллинг-Аллерёд. Этот генетический сдвиг показывает, что население Ближнего Востока, вероятно, уже начало перемещаться в Европу в конце верхнего палеолита, примерно на 6000 лет раньше, чем предполагалось ранее, до появления земледелия. Несколько экземпляров из кластера Виллабруна также показывают генетическое родство с восточными регионами, происходящими из потока генов. Вариант HERC2 для голубых глаз впервые появился около 13–14 тысяч лет назад в Италии и на Кавказе. светлая кожа пигментация, характерная для современных европейцев, по оценкам, распространилась по Европе "выборочно" во время мезолита (19-11 тыс. Лет назад). Связанные аллели TYRP1 SLC24A5 и SLC45A2 появляются около 19 тыс. Руб. Лет назад, еще во время LGM, скорее всего, на Кавказе.
 Упрощенная модель демографической истории европейцев в неолите период в внедрения сельского хозяйства
Упрощенная модель демографической истории европейцев в неолите период в внедрения сельского хозяйства 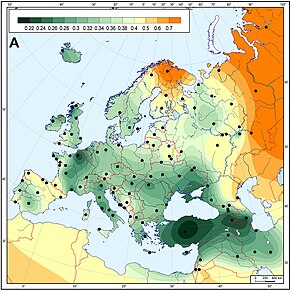 Древние европейские эпохи неолита были наиболее наиболее наиболее близки к современным ближневосточным / анатолийским популяциям. Генетические матрилинейные дистанции между популяциями европейской неолитической линейной керамической культуры (5 500–4900 до н.э.) и современными популяциями.
Древние европейские эпохи неолита были наиболее наиболее наиболее близки к современным ближневосточным / анатолийским популяциям. Генетические матрилинейные дистанции между популяциями европейской неолитической линейной керамической культуры (5 500–4900 до н.э.) и современными популяциями. Большая линия генетической изменчивости, которая давно признана в Европе, кажется, имеет важные отклонения от направления Срединного Восток. Это часто связывают с распространением сельскохозяйственных технологий в период неолита, который считается одним из самых важных периодов в определении современного европейского генетического разнообразия.
Неолит начался с распространения земледелия в юго-восточной Европе примерно в 10–3000 гг. До н. Э. И распространился на северо-запад Европы между 4500–1700 гг. До н. Э. В эту эпоху неолитическая революция привела к радикальным экономическим, а также социально-культурным изменениям в Европе, и считается, что это оказало большое влияние на генетическое разнообразие Европы, особенно в отношении генетических линий, попавших в Европу из Ближний Восток на Балканы. В этом периоде было несколько фаз:
Важный вопрос, касающийся генетического воздействия неолитических технологий в Европе - это способ, которым они были перенесены в Европу. Земледелие было введено значительной миграцией фермеров с Ближнего Востока (биологическая демическая диффузия модель Кавалли-Сфорца) или «культурной диффузией » или их комбинацией, и популяционные генетики пытались выяснить, соответствуют ли какие-либо генетические признаки ближневосточного происхождения маршрутам экспансии, постулированным археологическими свидетельствами.
Мартин Ричардс подсчитал, что только 11% европейской мтДНК является результатом иммиграции в Этот период свидетельствует о том, что земледелие было распространено в первуюочередь из-за того, что было принято коренным мезолитическим населением, а не из-за иммиграции с Ближнего Востока. Поток генов из Юго-Восточной Европы в Северо-Западную Европу, по-видимому, продолжался в неолите, и процентная доля значительно снижалась по направлению к Британским островам. Классическая генетика также предположила, что самая большая примесь к европейскому палеолиту / мезолиту была связана с неолитической революцией 7-5 тысячелетий до нашей эры. Были идентифицированы три группы генов мтДНК, внесшие вклад в развитие Европы в эпоху неолита: J, T1 и U3 (в указанном порядке важности). Вместе с другими они составляют около 20% генофонда .
В 2000 году исследование Semino на Y ДНК выявило наличие гаплотипов, принадлежащих к большой кладе E1b1b1 (E-M35). Они были преимущественно найдены на южных Балканах, юге Италии и некоторых частях Иберии. Семино связал этот паттерн, наряду с субкладами гаплогруппы J, как компонент Y-ДНК в неолитической демической диффузии фермеров с Ближнего Востока Кавалли-Сфорца. Россер и др. скорее рассматривает это как (прямой) «североафриканский компонент» в европейском генеалогии, хотя они не предлагают время и механизм для его учета. Андерхилл и Кивисилд (2007) ошибка harvcoltxt: нет цели: CITEREFUnderhill_and_Kivisild2007 (help ) также описал E1b1b как представляющий позднюю плейстоценовую миграцию из Африки через Синайский полуостров в Египта, что свидетельствует о том, что проявляется в митохондриальной ДНК.
 Современное распределение гаплогрупп Y-ДНК в каждой европейской стране
Современное распределение гаплогрупп Y-ДНК в каждой европейской стране Что касается распределения времени и разнообразия V13, Battaglia et al. (2008) ошибка harvcoltxt: нет цели: CITEREFBattaglia_et_al.2008 (help ) использует более раннее движение, в соответствии с соответствующей преемственности E-M78 *, являющаяся предком современных мужчин E-V13, быстро покинула южноегипетский родина и прибыла в Европу только с мезолитическими технологиями. Они затем предполагают, что подкласс E-V13 от E-M78 расширился только, когда коренные балканские «фуражиры-фермеры» перен технологии неолита с Ближнего Востока. Они предполагают, что первое крупное расселение E-V13 с Балкан могло произойти в направлении Адриатического моря с неолитической культурой импрессивной керамики, которую часто называют Impressa или Кардиал. Peričic et al. (2005) ошибка harvcoltxt: нет цели: CITEREFPeričic_et_al.2005 (help ), скорее предполагают, что основной маршрут распространения E-V13 пролегал по системе «шоссе» реки Вардар-Морава-Дунай.
В отличие от Battaglia, Cruciani et al. (2007) ошибка harvcoltxt: нет цели: CITEREFCruciani_et_al.2007 (help ) первоначальное (i) другая точка, где мутация V13 произошла на пути из Египта на Балканы через Ближний Восток, и (ii) более позднее время диспергирования. Авторы предположили, что мутация V13 впервые появилась в Интернационале Азии, где она проникла через Интернет примерно через 11 тыс. Руб. Лет назад. Позже он испытал быстрое распространение, которое он датировал ок. 5300 лет назад в Европе, совпало с эпохой балканской бронзы. Как Peričic et al. они считают, что «гаплогруппы E-V13 и J-M12 рассеялись, по-видимому, в основном по речным водным путям, соединяющим южные Балканы с северной и центральной Европой».
Совсем недавно Lacan et al. (2011) ошибка harvcoltxt: нет цели: CITEREFLacanKayserRicautBrucato2011 (help ) объявил, что 7000-летний скелет в контексте неолита в испанской погребальной пещере был человеком E-V13. (Другие образцы, протестированные на том же участке, относились к гаплогруппе G2a, которая была обнаружена в контексте неолита по всей Европе.) С помощью 7 STR-маркеров этот образец был идентифицирован как похожий на современных людей, протестированных в Албания, Босния, Греция, Корсика и Прованс. Поэтому авторы предположили, что независимо от того, является ли современное распространение E-V13 сегодня недавними недавними событиями, E-V13 уже находился в Европе в эпоху ранними ранними событиями и был перенесен фермерами из Восточного Средиземноморья в Западное Средиземноморье. намного раньше бронзового века. Это поддерживает предложения Battaglia et al. а не Cruciani et al. по крайней мере, в отношении самых ранних разгонов, но E-V13, возможно, разошелся более раза. Было высказано предположение, что современное распространение E-V13 в Европе даже позже, чем бронзовый век, по крайней мере частично вызвано перемещениями людей римской эпохи. (См. Ниже.)
. Миграция неолитических фермеров в Европу принесла с собой несколько новых приспособлений. Вариация светлого цвета кожи была представлена в Европе фермерами неолита. После достижения неолитических фермеров была выбрана мутация SLC22A4, что, вероятно, возникла для борьбы с дефицитом эрготионеина, но увеличивает риск язвенного колита, глютеновая болезнь и синдром раздраженного кишечника.
бронзовый век ознаменовался развитие междугородних торговых сетей, особенно вдоль Атлантического побережья и в долине Дуная. В этот период произошла миграция из Норвегии на Оркнейские острова и Шетландские (в меньшей степени в материковую Шотландию и Ирландию). Также была миграция из Германии в восточную Англию. Мартин Ричардс подсчитал, что в эпоху бронзы иммиграция мтДНК в Европу составляла около 4%.
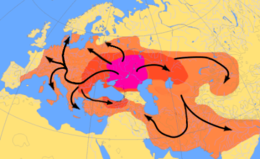 Схема индоевропейских миграций ок. С 4000 по 1000 г. до н.э., Курганской гипотезе
Схема индоевропейских миграций ок. С 4000 по 1000 г. до н.э., Курганской гипотезе Другая теория происхождения индоевропейского языка сосредоточена вокруг гипотетического приндоевропейского народа, Согласно гипотезе, курган может быть прослежен к северу от Черного и Каспийского морей примерно в 4500 году до нашей эры. Они приручили лошадь и, возможно, изобрели деревянный диск колесо. Считается, что они распространили свою культуру и гены по всей Европе. Гаплогруппа Y R1a является предполагаемым маркером этих "курганских" генов, как и гаплогруппа Y R1b, хотя эти гаплогруппы в целом могут быть намного старше языковой.
На крайнем севере носители Y-гаплогруппы N прибыли в Европу из Сибири, в конечном итоге расширившись до Финляндии, хотя конкретное время их получения неизвестно. Возраст наиболее распространенного субклада N1c1 в Северной Европе оценивается примерно в 8000 лет. Есть свидетельства поселения людей в Финляндии, датируемого 8500 г. до н.э., связанного с культурой Кунда и ее предполагаемым предком, культурой Свидера, но считается, что последняя имеет европейское происхождение.. Географическое распространение гаплогруппы N в Европе хорошо согласуется с культурой ямочно-гребенчатой керамики, появление которой обычно датируется ок. 4200 г. до н.э., и с распространением уральских языков. Исследования митохондриальной ДНК саамов, гаплогруппа U5 подтверждают множественные перемещения в Скандинавию из Волги - Урала
Отношения между странами европейских и азиатских колонистов в доисторические времена Финляндии являются предметом некоторых споров, и некоторые науки настаивают на том, что финны «преимущественно восточноевропейские и произведенные набор людей, которые отправились на север от украинского убежища во время ледникового периода». Дальше на восток вопрос менее спорный. Носители гаплогруппы составляют значительную часть всех неславянских этнических групп на севере России, в том числе 37% карелов, 35% коми (65% согласно другому исследованию), 67% марийцев, целых 98% ненцев, 94% нганасан, 86% - 94% якутов.
Ямный компонент содержит частичное происхождение от древнего североевразийского компонента, идентифицированного в Мальта. По словам Иосифа Лазаридиса, «древнее североевразийское происхождение является наименьшим компонентом повсюду в Европе, никогда не превышает 20 процентов, но мы находим его почти в каждой европейской группе, которую мы изучили». Этот генетический компонент линии происходит напрямую из самой передачи Мальта, а из родственной линии, которая отделилась от линии передачи Мальта.
До половины компонента Ямной может происходить из Кавказский охотник-собиратель прядь. 16 ноября 2015 года в исследовании, опубликованном в журнале Nature Communications, генетики объявили, что они представлены новое четвертое наследственное «племя» или «цепь», которая внесла свой вклад в современный европейский генофонд. Они проанализировали геномы двух охотников-собирателей из Грузии, которым было 13 300 и 9700 лет, и представили, что эти кавказские собиратели, вероятно, были получены ДНК фермеров в Ямне. По словам соавтора доктора Андреа Маника из Кембриджского университета: «Вопрос о том, откуда взялись Ямная, до сих пор оставался загадкой... теперь мы можем ответить на этот вопрос, как мы видим, что их генетическое происхождение. -up представляет собой смесь охотников-собирателей из Восточной Европы и населения из этого кармана кавказских охотников-собирателей, которые пережили большую часть последнего ледникового периода в кажущейся изоляции. "
Согласно Лазаридису и другие. (2016), население, относящееся к людям хальколита Ирана, составляющая примерно половину предков ямных популяций Причерноморско-Каспийской степи. Эпохи энеолита были смесью «неолитических людей западного Ирана, Леванта и кавказских охотников-собирателей».
Генетические вариации стойкости лактазы и большего роста пришли с ямной людьми.Производный аллельена KITLG (SNP rs12821256), который связан с светлыми волосами и может быть причиной - светлых волос у европейцев, обнаружен в популяциях с восточными, но не с западными охотниками. Предки собирателей, что позволяет предположить, что его происхождение относится к населению Древней Северной Евразии (ANE) и, возможно, было распространено в Европе людьми с степным происхождением. этим, самым ранним научным человеком с производным аллелем является особь ANE из археологи ческого комплекса позднего верхнего палеолита Афонтова гора в центральной Сибири.
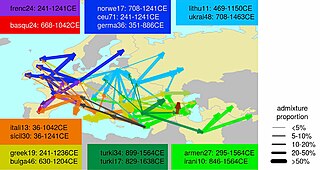 Обзорная карта недавнего времени (1–17 вв. Н.э.) примесные события в Европе
Обзорная карта недавнего времени (1–17 вв. Н.э.) примесные события в Европе В период Римской империи исторические источники показывают, что было много перемещений людей по Европе, как внутри, так и за пределами Империи. Иногда упоминаются случаи геноцида, совершенного римлянами против мятежных провинциальных племен. Если бы это действительно произошло, это было бы ограниченным, учитывая, что современные популяции демонстрируют значительную генетическую преемственность в своих соответствующих регионах. Процесс «романизации », по-видимому, завершился колонизацией провинций несколькими латинскими администраторами, военнослужащими, оседлыми ветеранами и частными лицами (купцами, торговцами), которые происходили из различных регионов Империи (а не только из Римской Италии ). Они послужили ядром для аккультурации местной знати.
Учитывая их небольшую численность и различное происхождение, романизация, похоже, не оставила отчетливых генетических признаков в Европе. Действительно, было обнаружено, что романоязычные народы на Балканах, такие как румыны, армыны, молдаване и т. Д., Генетически напоминают соседних греков и южнославяноязычных. людей, а не современных итальянцев, доказывая, что они были генетически говорящими, в основном через I2a2 M-423 и E1b1b1, V-13 гаплогруппы коренных жителей этой области.
Стивен Берд предположил, что E1b1b1a распространился в период римской эпохи через фракийское и дакское население с Балкан в остальную Европу.
позднеримский период (не только) германского «Völkerwanderung », были сделаны некоторые предположения, по крайней мере для Британии, с гаплогруппой Y1a, связанной с англосаксонской иммиграция в восточную Англию, и R1a связан с норвежской иммиграцией в северной Шотландии.
Есть f наша основная ДНК Y-хромосомы гаплогруппы, которые составляют большую часть отцовского происхождения в Европе.
Помимо небольших анклавов, есть также несколько гаплог, помимо вышеуказанных четырех, которые являются наиболее распространены в определенных регионах. Европы.
В Европе был проведен ряд исследований по гаплогруппам митохондриальной ДНК (мтДНК). В отличие от гаплогрупп ДНК Y, гаплогруппы мтДНК не демонстрировали такой большой географической структуры, но были более равномерно распространены. За исключением отдаленных саамов, для всех европейских характерно преобладание гаплогрупп H, U и T. Отсутствие наблюдаемой географической структуризации мтДНК может быть связано с социокультурными факторами, а именно явлениями полигинии и патрилокальность.
Генетические предполагаемые исследования некоторый поток материнских генов в Восточную Европу из Восточной Азии или южной Сибири 13 000–6600 лет л.н.. Анализ неолитических скелетов на Великой Венгерской равнине какой великой Венгерской равнине, некоторые из демонстрируют выживают в современных популяциях Восточной Европы. Поток материнских генов в Европу из Африки к югу от Сахары начался уже 11000 лет назад, хотя, по оценкам большинства ген родословных, примерно 65%, появилось позже, в том числе в период романизации, арабских завоеваний южной Европы и во время работорговли в Атлантике.
Исследование 2007 года, проведенное Bauchet, в котором использовалось около 10 000 аутосомных SNP ДНК, дало аналогичные результаты. Основные компоненты выявил четыре широко рассредоточенные группы, соответствующие Африке, Европе, Центральной Азии и Южной Азии. PC1 отделял африканцев от других популяций, PC2 отделял азиатов от европейцев и африканцев, в то время как PC3 отделял жителей Центральной Азии от жителей Южной Азии.
| Северо-Западная Европа (CEU) | Йоруба | Хань китайский | |
|---|---|---|---|
Северо-Западная Европа (CEU) | 0,1530 | 0,1100 | |
| Йоруба | 0,1530 | 0,1900 | |
| Хань китайский | 0,1100 | 0,1900 |
| Европа | Америка | Восточная Азия | Океания | |
|---|---|---|---|---|
| Америка | 9,5 | |||
| Восточная Азия | 9,7 | 8,9 | ||
| Океания | 13,5 | 14,6 | 10 | |
| Африка | 16,6 | 22,6 | 20,6 | 24,7 |
Генетически Европа относительно однородна, но отличаются -популяционные паттерны типов генетических маркеров имеют обнаружены, особенно вдоль склона юго-восток-северо-запад. Например, анализ основных компонентов Кавалли-Сфорца выявил пять основных клинических паттернов по всей Европе, и аналогичные паттерны продолжали обнаруживаться в более поздних исследованиях.
Он также создал филогенетическое древо для анализа внутренних отношений между европейцами. Он обнаружил четырех основных «исключений»: басков, саамов, сардинцев и исландцев ; результат он объяснил их относительной изоляцией (примечание: исландцы и сардинцы говорят на индоевропейских языках, а две другие группы - нет). греки и югославы представляли вторую группу менее радикальных исключений. Остальные популяции сгруппированы в несколько групп: «кельтский », «германский », «юго-западные европейцы», «скандинавы » и «восточноевропейцы».
Исследование 19 популяций из Европы, проведенное в мае 2009 г. с использованием 270 000 SNP, выявило генетическое разнообразие европейских популяций, соответствующее градиенту с северо-запада на юго-восток, и выделило «отдельных регионов» в Европе:
В этом исследовании анализ барьеров показал «генетические барьеры» между Финляндией, Италией и другими странами, и эти барьеры также могут быть в Финляндии (между Хельсинки и Куусамо) и Италии (между северной и южной частями, Fst = 0,0050). Было обнаружено, что Fst (Индекс фиксации ) значительно коррелирует с географическими расстояниями в диапазоне от ≤0,0010 для соседних популяций до 0,0200–0,0230 для южной Италии и Финляндии. Для сравнения, попарные Fst неевропейских выборок составили: европейцы - африканцы (йоруба) 0,1530; Европейцы - китайцы 0,1100; Африканцы (йоруба) - китайцы 0,1900.
Исследование, проведенное Чао Тианом в августе 2009 г., расширило анализ генетической структуры европейской популяции, включив в нее дополнительные южноевропейские группы и арабское население (палестинцы, Друзы...) с Ближнего Востока. Это исследование определило аутосомный Fst между 18 группами населения и пришло к выводу, что в целом генетические отличия соответствуют географическим отношениям с меньшими значениями между группами населения с происхождением изних стран / регионов (, греки / Тосканцы : Fst = 0,0010, греки / палестинцы : Fst = 0,0057) по сравнению с жителями очень разных регионов Европы (например, греки / Шведский : Fst = 0,0087, Греки / Русские : Fst = 0,0108).
Селдин (2006) использовал более 5000 аутосомных SNP. Он показал «последовательное и воспроизводимое различие между« северными »и« южными »группой европейского населения ». Большинство участников южноевропейского (итальянцы, греки, португальцы, испанцы ) и Евреи- ашкенази составляют>85% южного населения; и большинство северных, западных, центральных и восточных европейцев (шведов, англичан, ирландцев, немцев и украинцев ) имеют>90% в северной группе населения. Однако многие из участников этого рынка самом деле были идентифицированы себя американскими гражданами, которые идентифицировали себя с разными европейскими этническими группами на основании собственной семейной родословной.
Аналогичное исследование, проведенное в 2007 году с использованием выбора исключительно из Европы, показало наиболее важную генетическую дифференциацию в Европе по линии с севера на юго-восток (северная Европа на Балканы), с другой осью дифференциации восток-запад через Европу. Его результаты согласились с более ранними результатами, основанными на мтДНК и Y-хромосомной ДНК, которые подтверждают теорию о том, что современные иберийцы (испанские и португальские) имеют самую древнюю европейскую генетическую родословную, а также отделяют басков и саамов от других популяций.
Предполагается, что английский ирландский кластер с другими северными и восточными европейцами, такими как немцы и поляки, в то время как некоторые баски и итальянцы также сгруппировались с Северные европейцы. Несмотря на эти стратификации, он отличается высокой степенью однородности в Европе и на Ближнем Востоке: «в Европе наблюдается низкое видимое разнообразие, при этом выборки в масштабах всего континента незначительно более разбросаны, чем отдельные выборки популяций в других странах мира». такие как балканские народы (такие как фригийцы и македонские греки ),
В 2008 году две международные исследовательские группы опубликовали анализ крупномасштабного генотипирования больших выборок европейцев, используя более 300 000 аутосомных SNP. За исключением обычных изолятов, таких как басков, финнов и сардинцев, в европейской популяции отсутствовали резкие разрывы (кластеризация), как показали предыдущие исследования (см. Селдин) et al., 2006 и Bauchet et al. 2007), хотя был заметный градиент с юга на север. В целом они представлены лишь низким уровнем генетической дифференциации между субпопуляциями, которые действительно существовали, характеризовались сильным континентальная корреляция между географическими и географическими расстояниями. Кроме того, они представлены, что разнообразие было самым большим в южной Европе из-за более крупного эффективного размера популяции и / или экспансии населения с юга на север Е вропы. Исследователи предполагают подразумевает отдельные европейцы не делятся на популяции.
Исследование северо-восточных популяций, опубликованное в марте 2013 года, показало, что народы коми сформировали полюс генетического разнообразия, отличного от других популяции.
Все современные европейцы и жители Ближнего Востока обладают необычно высокой степенью генетической однородности в Европе и на Ближнем Востоке. такие как балканские народы (такие как фригийцы и македонские греки ),
Все современные европейцы и современные жители Ближнего Востока имеют необычно высокую степень общего генетического наследия предков.>балканские народы (такие как фригийцы и македонские греки ),
.
генетическая дистанция между популяциями часто измеряется с С помощью индекса фиксации (Fst), основанного на данных генетического полиморфизма, таких как или микросателлиты, таких как или микросателлиты. Fst - это особый случай F -статистики, концепция, разработанная в 1920-х годах Сьюэллом Райтом. Fst - это просто корреляция случайно выбранных аллелей в одной и той же подгруппе населения относительно аллелей, обнаруженных во всем Часто выражается как доля генетического разнообразия из-за различий в частотах аллелей среди популяций.
Диапазон значений от 0 до 1. Нулевое значение подразумевает, что эти две популяции являются панмиктическими, что они свободно скрещиваются. Значение будет означать, что две популяции полностью разделены. Чем больше значение Fst, тем больше генетическая дистанция. По сути, эти низкие значения Fst предполагают, что большая часть генетической изменчивости находится на уровне индивидов в одной и той же группе населения (~ 85%); принадлежность к разным группам населения в пределах одной «расы» / континента и даже к разным расовым / континентальным группам добавляла меньшую гораздо большую степень вариации (3–8%; 6–11%, соответственно).
| итальянские американцы | палестинцы | шведы | друзы | испанцы | немцы | русские | ирландцы | греко-американцы | евреи ашкенази | черкесы | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| итальянские американцы | 0,0064 | 0, 0064 | 0,0057 | 0,0010 | 0,0029 | 0,0088 | 0,0048 | 0,0000 | 0,0040 | 0,0067 | |
| Палестинцы | 0,0064 | 0,0191 | 0, 0064 | 0,0101 | 0,0136 | 0,0202 | 0,0170 | 0,0057 | 0,0093 | 0,0108 | |
| шведы | 0,0064 | 0,0191 | 0,0167 | 0, 0040 | 0,0007 | 0,0030 | 0,0020 | 0,0084 | 0,0120 | 0,0117 | |
| Друзы | 0,0057 | 0,0064 | 0,0167 | 0,0096 | 0, 0121 | 0,01 94 | 0,0154 | 0,0052 | 0,0088 | 0,0092 | |
| Испанцы | 0,0010 | 0,0101 | 0,0040 | 0,0096 | 0,0015 | 0,0070 | 0, 0037 | 0,0035 | 0,0056 | 0,0090 | |
| Немцы | 0,0029 | 0,0136 | 0,0007 | 0,0121 | 0,0015 | 0,0030 | 0,0010 | 0, 0039 | 0,0072 | 0,0089 | |
| Русские | 0,0088 | 0,0202 | 0,0030 | 0,0194 | 0,0070 | 0,0030 | 0,0038 | 0,0108 | 0, 0137 | 0,0120 | |
| Ирландский | 0,0048 | 0,0170 | 0,0020 | 0,0154 | 0,0037 | 0,0010 | 0,0038 | 0,0067 | 0,0109 | 0, 0110 | |
| Американцы греческого происхождения | 0,0000 | 0,0057 | 0,0084 | 0,0052 | 0, 0035 | 0,0039 | 0,0108 | 0,0067 | 0,0042 | 0,0054 | |
| евреи ашкенази | 0,0040 | 0,0093 | 0,0 120 | 0,0088 | 0,0056 | 0,0072 | 0,0137 | 0,0109 | 0,0042 | 0,0107 | |
| Черкесы | 0,0067 | 0,0108 | 0,0117 | 0,0092 | 0,0090 | 0,0089 | 0,0120 | 0,0110 | 0,0054 | 0,0107 |
| Австрия | Болгария | Чешская Республика | Эстония | Финляндия (Хельсинки ) | Финляндия ( Куусамо ) | Франция | Северная Германия | Южная Германия | Венгрия | Северная Италия | Южная Италия | Латвия | Литва | Польша | Россия | Испания | Швеция | Швейцария | CEU | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Австрия | 1,14 | 1,08 | 1,58 | 2, 24 | 3,30 | 1,16 | 1,10 | 1,04 | 1,04 | 1,49 | 1,79 | 1,85 | 1,70 | 1,19 | 1,47 | 1,41 | 1,21 | 1,19 | 1,12 | Австрия | |
| Болгария | 1,14 | 1,21 | 1,70 | 2,19 | 2,91 | 1,22 | 1,32 | 1,19 | 1,10 | 1,32 | 1,38 | 1,86 | 1,73 | 1,29 | 1,53 | 1,30 | 1,47 | 1,13 | 1,29 | Болгария | |
| Чешская Республика | 1,08 | 1,21 | 1,42 | 2,20 | 3,26 | 1,35 | 1,15 | 1,16 | 1,06 | 1,69 | 2,04 | 1,62 | 1,48 | 1,09 | 1,27 | 1,63 | 1,26 | 1,37 | 1,21 | Чешская Республика | |
| Эстония | 1,58 | 1,70 | 1,42 | 1,71 | 2,80 | 2,08 | 1,53 | 1,70 | 1,41 | 2,42 | 2,93 | 1,24 | 1,28 | 1,17 | 1,21 | 2,54 | 1,49 | 2,16 | 1,59 | Эстония | |
| Финляндия (Хельсинки) | 2,24 | 2,19 | 2,20 | 1,71 | 1,86 | 2,69 | 2,17 | 2,35 | 1,87 | 2,82 | 3,37 | 2,31 | 2,33 | 1,75 | 2,10 | 3,14 | 1,89 | 2,77 | 1,99 | Финл яндия (Хельсинки) | |
| Финляндия (Куусамо) | 3,30 | 2,91 | 3,26 | 2,80 | 1,86 | 3,72 | 3,27 | 3,46 | 2,68 | 3,64 | 4,18 | 3,33 | 3,37 | 2,49 | 3,16 | 4,21 | 2,87 | 3,83 | 2,89 | Финляндия (Куусамо) | |
| Франция | 1,16 | 1,22 | 1,35 | 2,08 | 2,69 | 3,72 | 1,25 | 1,12 | 1,16 | 1,38 | 1,68 | 2,40 | 2,20 | 1,44 | 1,94 | 1,13 | 1,38 | 1,10 | 1,13 | Франция | |
| Северная Германия | 1,10 | 1,32 | 1,15 | 1,53 | 2,17 | 3,27 | 1,25 | 1,08 | 1,11 | 1,72 | 2,14 | 1,84 | 1,66 | 1,18 | 1,49 | 1,62 | 1,12 | 1,36 | 1,06 | Северная Германи я | |
| Южная Германия | 1,04 | 1,19 | 1,16 | 1,70 | 2,35 | 3,46 | 1,12 | 1,08 | 1,08 | 1,53 | 1,85 | 1,20 | 1,84 | 1,23 | 1,58 | 1,40 | 1,21 | 1,17 | 1,07 | Южная Германия | |
| Венгрия | 1,04 | 1,10 | 1,06 | 1,41 | 1,87 | 2,68 | 1,16 | 1,11 | 1,08 | 1,42 | 1,63 | 1,58 | 1,46 | 1,14 | 1,28 | 1,32 | 1,22 | 1,16 | 1,13 | Венгрия | |
| Северная Италия | 1,49 | 1,32 | 1,69 | 2,42 | 2,82 | 3,64 | 1,38 | 1,72 | 1,53 | 1,42 | 1,54 | 2,64 | 2,48 | 1,75 | 2,24 | 1,42 | 1,86 | 1,36 | 1,56 | Северная Италия | |
| Южная Италия | 1, 79 | 1,38 | 2,04 | 2,93 | 3,37 | 4,18 | 1,68 | 2,14 | 1,85 | 1,63 | 1,54 | 3,14 | 2,96 | 1,99 | 2,68 | 1,67 | 2,28 | 1,54 | 1,84 | Южная Италия | |
| Латвия | 1,85 | 1,86 | 1,62 | 1,24 | 2,31 | 3,33 | 2,40 | 1,84 | 1,20 | 1,58 | 2,64 | 3,14 | 1,20 | 1,26 | 1,84 | 2,82 | 1,89 | 2,52 | 1,87 | Латвия | |
| Литва | 1,70 | 1,73 | 1,48 | 1,28 | 2,33 | 3,37 | 2,20 | 1,66 | 1,84 | 1,46 | 2,48 | 2,96 | 1,20 | 1,20 | 1,26 | 2,62 | 1,74 | 2,29 | 1,74 | Литва | |
| Польша | 1,19 | 1,29 | 1,09 | 1,17 | 1,75 | 2,49 | 1,44 | 1,18 | 1,23 | 1,14 | 1,75 | 1,99 | 1,26 | 1,20 | 1,18 | 1,66 | 1,30 | 1,46 | 1,28 | Польша | |
| Россия | 1,47 | 1,53 | 1,27 | 1,21 | 2,10 | 3,16 | 1,94 | 1,49 | 1,58 | 1,28 | 2,24 | 2,68 | 1,84 | 1,26 | 1,18 | 2,32 | 1,59 | 1,20 | 1,56 | Россия | |
| Испания | 1,41 | 1,30 | 1,63 | 2,54 | 3,14 | 4,21 | 1,13 | 1,62 | 1,40 | 1,32 | 1,42 | 1,67 | 2,82 | 2,62 | 1,66 | 2,32 | 1,73 | 1,16 | 1,34 | Испания | |
| Швеция | 1,21 | 1,47 | 1,26 | 1,49 | 1,89 | 2,87 | 1,38 | 1,12 | 1,21 | 1,22 | 1,86 | 2,28 | 1,89 | 1,74 | 1,30 | 1,59 | 1,73 | 1,50 | 1,09 | Швеция | |
| Швейцария | 1,19 | 1,13 | 1,37 | 2,16 | 2,77 | 3,83 | 1,10 | 1,36 | 1,17 | 1,16 | 1,36 | 1,54 | 2,52 | 2,29 | 1,46 | 1,20 | 1,16 | 1,50 | 1,21 | Швейцария | |
| CEU | 1,12 | 1,29 | 1,21 | 1,59 | 1,99 | 2,89 | 1,13 | 1,06 | 1,07 | 1,13 | 1,56 | 1,84 | 1,87 | 1,74 | 1,28 | 1,56 | 1,34 | 1,09 | 1,21 | CEU | |
| Австрия | Болгария | Чехия | Эстония | Финляндия (Хельсинки) | Финляндия (Куусамо) | Франция | Северная Германия | Южная Германия | Венгрия | Северная Италия | Южная Италия | Латвия | Литва | Польша | Россия | Испания | Швеция | Швейцария | CEU |
Одним из первых ученых, выполнивших генетические исследования, был Луиджи Лука Кавалли-Сфорца. Он использовал классические генетические маркеры для прокси-анализа ДНК. Этот метод изучает различия в частотах конкретных аллельных признаков, а именно полиморфизмов белков, обнаруженных в крови человека (таких как группы крови ABO, антигены крови резуса, локусы HLA, иммуноглобулины, G6PD изоферменты и другие). Впоследствии его команда вычислила генетическое расстояние между популяциями, исходя из принципа, что две популяции с одинаковыми частотами признака более тесно связаны, чем популяции с более разными частотами признака.
Исходя из этого, он построил филогенетические деревья, которые схематически отображали генетические расстояния. Его команда также выполнила анализ главных компонентов, который хорош для анализа многомерных данных с минимальной потерей информации. Утерянную информацию можно частично восстановить, создав второй главный компонент и так далее. В свою очередь, информация от каждого отдельного главного компонента (PC ) может быть представлена графически на синтетических картах. На этих картах показаны пики и впадины, которые представляют популяции, чьи частоты генов принимают экстремальные значения по сравнению с другими в исследуемой области.
Пики и впадины, обычно соединенные плавными градиентами, называются линиями. Генетические клины могут быть созданы путем адаптации к окружающей среде (естественный отбор ), непрерывного потока генов между двумя изначально разными популяциями или демографической экспансии в малонаселенную среду с небольшим начальным примесь с существующими популяциями. Кавалли-Сфорца связал эти градиенты с постулируемыми доисторическими движениями населения, основанными на археологических и лингвистических теориях. Однако, учитывая, что временная глубина таких закономерностей неизвестна, «связывание их с конкретными демографическими событиями обычно является умозрительным».
Исследования с использованием прямого анализа ДНК сейчас многочисленны и может использовать митохондриальную ДНК (мтДНК), нерекомбинирующую часть Y-хромосомы (NRY) или даже аутосомную ДНК. ДНК MtDNA и NRY имеют некоторые общие черты, которые сделали их особенно полезными в генетической антропологии. Эти свойства включают прямое неизмененное наследование мтДНК и ДНК NRY от матери к потомству и от отца к сыну, соответственно, без «скремблирующих» эффектов генетической рекомбинации. Мы также предполагаем, что эти генетические локусы не подвержены естественному отбору и что основным процессом, ответственным за изменения пар оснований, была мутация (которую можно вычислить).
Меньший эффективный размер популяции NRY и мтДНК усиливает последствия дрейфа и эффекта основателя по сравнению с аутосомами, что делает вариацию NRY и мтДНК потенциально чувствительным показателем состава популяции. Эти биологически правдоподобные предположения не являются конкретными; Россер предполагает, что климатические условия могут влиять на плодовитость определенных линий.
Основная частота мутаций, используемая генетиками, является более сомнительной. Они часто используют разную частоту мутаций, и исследования часто приходят к совершенно разным выводам. NRY и мтДНК могут быть настолько восприимчивы к дрейфу, что некоторые древние паттерны могут быть скрыты. Другое предположение состоит в том, что генеалогии населения аппроксимируются генеалогиями аллелей . Гвидо Барбуджани указывает, что это справедливо только в том случае, если группы населения развиваются из генетически мономорфного набора основателей. Барбужани утверждает, что нет оснований полагать, что Европа была колонизирована мономорфными популяциями. Это привело бы к переоценке возраста гаплогруппы, что привело бы к ошибочному расширению демографической истории Европы до позднего палеолита, а не неолита. Большую уверенность в хронологии можно получить из исследований древней ДНК (см. Ниже), но до сих пор их было сравнительно немного.
В то время как гаплогруппы Y-ДНК и мтДНК представляют собой лишь небольшой компонент пула ДНК человека, аутосомная ДНК имеет то преимущество, что содержит сотни тысяч исследуемых генетических локусов., давая более полную картину генетического состава. Родственные отношения могут быть определены только на статистической основе, потому что аутосомная ДНК подвергается рекомбинации. Одна хромосома может записывать историю каждого гена. Аутосомные исследования гораздо более надежны для демонстрации взаимоотношений между существующими популяциями, но не дают возможности раскрыть их историю так, как обещают исследования мтДНК и ДНК NRY, несмотря на их многочисленные сложности.
Генетические исследования основаны на многочисленных предположениях и страдают методологическими ограничениями, такими как систематическая ошибка отбора и смешанные явления, такие как генетический дрейф, фундаментальные эффекты и эффекты узкого места вызывают большие ошибки, особенно в исследованиях гаплогруппы. Независимо от того, насколько точна методология, выводы, сделанные на основе таких исследований, основываются на том, как автор рассматривает свои данные в соответствии с установленными археологическими или лингвистическими теориями.
| На Wikimedia Commons есть материалы, связанные с генетическими исследованиями европейских популяций. |
.