
Войти
Газообмен - это физический процесс, посредством которого газы движутся пассивно посредством диффузии по поверхности. Например, эта поверхность может быть границей раздела воздух / вода в водоеме, поверхность пузырька газа в жидкости, газопроницаемой мембраной или мембраной, образует границу между организмом и его внеклеточной средой.
Газы постоянно потребляются и вырабатываются эффективными системами газообмена между живыми, поэтому эффективная система газообмена между, в конечном счете, внутренним пространством клетки (-ей) и внешняя среда не требуется. Небольшие, особенно одноклеточные организмы, такие как бактерии и простейшие, имеют высокое отношение площади поверхности к объему. У этих существ есть газообменная мембрана - это обычно клеточная мембрана. Некоторые мелкие многоклеточные организмы, такие как плоские черви, также обладают достаточным газообменом через кожу или кутикулу, я их тела. Однако для более крупных организмов, которые имеют небольшое отношение площади поверхности к объему, есть специализированные структуры с извилистой поверхностью, такие как жабры, легочные альвеолы и мезофилл обеспечить большую площадь, используемое для эффективного газообмена. Эти извилистые поверхности иногда могут проникать внутрь организма. Так обстоит дело с альвеолами, которые образуют внутреннюю поверхность легких млекопитающих, губчатый мезофилл, который находится внутри листьев некоторых видов растений, или жабры тех моллюсков, у которых они есть, которые находятся в полости мантии.
У аэробных организмов газообмен особенно важен для дыхания, которое включает поглощение кислорода (O. 2) и выделение углерода. диоксид (CO. 2). И наоборот, в кислородных фотосинтезирующих организмах, таких как большинство наземных растений, поглощение углекислого газа и выделение как кислорода, так и водяного пара осуществляющих процессы газообмена, происходящих в течение дня. Другие процессы газообмена важны для менее известных организмов: например, диоксид углерода, метан и водород обмениваются через клеточную мембрану метаногенных архей. В фиксация диазотрофными бактериями и денитрификация гетеротрофными бактериями (такими как Paracoccus denitrificans и различные псевдомонады ), газообразный азот обменивается с окружающей средой, поглощается первым и выделяется в то время как гигантские трубчатые черви полагаются на бактерии, чтобы окислить сероводород, извлеченный из морской среды, используя растворенный в воде кислород в качестве акцептора электронов.
Обмен газов происходит в результате диффузии вниз по градиенту концентрации. Молекулы газа перемещаются в своей области, в которой они находятся в высокой концентрации. Диффузия - это пассивный процесс, означающий, что для питания не требуется энергия, и он следует закону Фика :

По отношению к типичной биологической системе, где два отсека («внутри» и «снаружи») разделены мембранным барьером, и где газу разрешается самопроизвольно выходить диффузия вниз по градиенту концентрации:

Рис. 1 Закон Фика для поверхности газообмена
Газы должны сначала раствориться в жидкости, чтобы диффундировать через мембрану, поэтому для всех биологических газообменных систем требуется влажная среда. Как правило, чем выше скорость концентрации на газообменной поверхности, тем выше скорость диффузии через нее. И наоборот, чем тоньше газообменная поверхность (при той же разнице концентраций), тем быстрее газы будут диффундировать через нее.
В приведенном выше уравнении J - это поток, выраженный на единицу площади, поэтому увеличение площади не повлияет на ее значение. Однако увеличение доступной площади поверхности увеличит количество газа, которое может диффундировать за заданное время. Это связано с тем, что количество газа, диффундирующего в единицу времени (dq / dt), является произведением J и площади газообменной поверхности, A:

одноклеточные организмы, такие как бактерии и амебы, не имеют поверхностей газообмена, потому что они могут использовать преимущества большой площади поверхности они имеют относительно их объема. Количество газа, которое производит (или требует) в данный момент времени, будет примерно пропорционально объему его цитоплазмы. Объем одноклеточного организма очень мал, поэтому он производит (и требует) относительно небольшое количество газа в данный момент времени. По сравнению с этим небольшим объемом клеточной поверхности его очень велика мембраны очень велика и без дополнительных вариантов удовлетворения потребностей в газообмене. Однако по мере увеличения размера организма его площадь и объем изменяются по-разному. Рассмотрим воображаемый организм, который представляет собой куба со стороной L. Его объем с размером поверхности увеличивается только на квадрат (L) его размер. Это означает, что внешняя поверхность становится непригодной для быстро развивающихся потребителей в газообмене большего объема цитоплазмы. Кроме того, толщина поверхности, которую газы должны иметь толщину (dx в законе Фика), также может быть больше у более тонких организмов: в случае одноклеточного организма типичная клеточная мембрана имеет толщину всего 10 нм; но у более тонких организмов, таких как круглые черви (Nematoda), эквивалентная поверхность обмена - кутикула - значительно толще и составляет 0,5 мкм.
 Рис. 2. Сравнение операций и эффектов системы прямотока и противоточной системы обмена показано на верхней и нижней диаграммах соответственно. В обоих случаях (и указывается), что красный цвет имеет более высокое значение (например, температура или парциального давления газа), чем синий, и что свойство, передаваемое в каналх, поэтому переходит от красного к синему. Обратите внимание, что каналы являются эффективными, если между ними происходит эффективный обмен (т. Е. Между каналами).
Рис. 2. Сравнение операций и эффектов системы прямотока и противоточной системы обмена показано на верхней и нижней диаграммах соответственно. В обоих случаях (и указывается), что красный цвет имеет более высокое значение (например, температура или парциального давления газа), чем синий, и что свойство, передаваемое в каналх, поэтому переходит от красного к синему. Обратите внимание, что каналы являются эффективными, если между ними происходит эффективный обмен (т. Е. Между каналами). Таким образом, у многоклеточных организмов для обеспечения организма часто используются используемые органы дыхания, такие как жабры или легкие. дополнительной площади поверхности для необходимой скорости газообмена с внешней средой. Однако расстояния между газообменником и более глубокими тканями слишком велики для удовлетворения потребностей этих тканей в газе. Поэтому газообменники часто присоединяются к газораспределительным системам кровообращения, которые транспортируют газы равномерно ко всем тканям независимо от их расстояния от газообменника.
Некоторые многоклеточные организмы, такие как плоские черви (Platyhelminthes) относительно большие, но очень тонкие, что позволяет их внешней поверхности тела действовать как поверхность газообмена без необходимости в специализированном газообменном органе. Поэтому у плоских червей отсутствуют жабры или легкие, а также отсутствует система кровообращения. Другие многоклеточные организмы, такие как губки (Porifera), по своей природе большую площадь поверхности, поскольку они очень пористые и / или разветвленные. Губкам не нужна кровеносная система или специализированные органы газообмена, потому что их стратегия питания включает одностороннюю прокачку воды через их пористые тела с использованием флагилированных воротниковых клеток. Таким образом, каждая клетка тела губки подвергается постоянному воздействию свежей насыщенной кислородом воды. Таким образом, они могут распространяться на диффузию через клеточные мембраны для осуществления газообмена, необходимого для дыхания.
У организмов, кровеносные системы, которые связаны с их специализированными газообменными поверхностями, для взаимодействия между ними.
В противоточной системе воздух (или, чаще всего, вода, содержащая растворенный воздух) втягивается в направлении, противоположном потоку крови в газообменнике. Противоточная система, такая как эта, поддерживает крутой градиент по длине газообменной поверхности (см. Нижнюю диаграмму на рис. 2). Так обстоит дело с жабрами рыб и разными водными существами. Газосодержащая вода из окружающей среды втягивается в одном направлении через поверхность газообмена, кровоток в нижних жаберных капиллярах течет в противоположном направлении. Хотя это теоретически позволяет почти полностью переносить респираторный газ от одной стороны теплообменника к другому, у рыб менее 80% кислорода в воде, протекающей по жабрам, обычно переносится в крови.
Альтернативные схемы системы перекрестного тока встречаются у птиц. и тупиковые системы мешков, заполненных воздухом, обнаруженные в легких млекопитающих. В системе с прямоточным потоком кровь и газ (или жидкость, содержащаяся газ) движутся в одном и том же направлении через газообменник. Это означает, что величина градиента меняется по длине поверхности газообмена, и в конечном итоге прекращается, когда будет достигнуто равновесие (см. Верхнюю диаграмму на рис. 2). Системы газообмена с прямоточным потоком, как известно, не используются в природе.
 Рис. 3. Альвеола (множественное число: альвеолы, от латинского альвеуса, «маленькая полость») - это анатомическая структура, имеющая форму полой полости. Они встречаются в легких млекопитающих. Они представляют собой сферические выходы респираторных бронхиол и респираторные органы, оказывающие воздействие на газообмена с кровью.
Рис. 3. Альвеола (множественное число: альвеолы, от латинского альвеуса, «маленькая полость») - это анатомическая структура, имеющая форму полой полости. Они встречаются в легких млекопитающих. Они представляют собой сферические выходы респираторных бронхиол и респираторные органы, оказывающие воздействие на газообмена с кровью. У млекопитающих газообменник интернализируется, образуя легкие, как и у представителей наземных животных. Газообмен происходит в микроскопических тупиковых мешочках, заполненных воздухом, называемых альвеолами, где очень тонкая мембрана (называемая гемато-воздушный барьер ) разделяет кровь в альвеолярных капиллярах (в стенке альвеол) из альвеолярного воздуха в мешочках.
 Рис. 4. Гистологический разрез альвеолярной стенки, показывающие слои, через которые газы перемещаются между плазмой и альвеолярным воздухом. Темно-синие объекты представляют собой ядра капиллярных эндотелиальных и альвеолярных клеток типа I эпителия (или типа 1 пневмоцитов ). Два красных объекта с надписью «РБК» - это эритроциты в крови альвеолярных капилляров.
Рис. 4. Гистологический разрез альвеолярной стенки, показывающие слои, через которые газы перемещаются между плазмой и альвеолярным воздухом. Темно-синие объекты представляют собой ядра капиллярных эндотелиальных и альвеолярных клеток типа I эпителия (или типа 1 пневмоцитов ). Два красных объекта с надписью «РБК» - это эритроциты в крови альвеолярных капилляров. Мембрана, через которую происходит газообмен в альвеолах (т. Е. Гемато-воздушный барьер), очень тонкая (у человека в среднем толщиной 2,2 мкм). Он состоит из альвеолярных эпителиальных клеток, их базальных мембран и эндотелиальных клеток легочных капилляров (рис. 4). Большая площадь поверхности мембраны обусловлена складыванием мембраны примерно в 300 миллионов альвеол с диаметром примерно 75-300 мкм каждую. Это обеспечивает большую площадь поверхности (145 м), на которой может происходить газообмен.
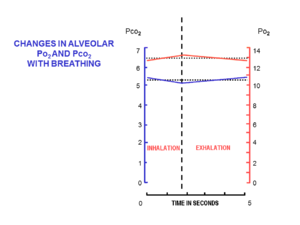 Рис. 5. Изменения в составе альвеолярного воздуха во время нормального дыхательного цикла в состоянии покоя. Шкала слева и синяя линия указывает парциальные давления углекислого газа в кПа, а шкала справа и красная линия указывает парциальные давления кислорода, также в кПа (чтобы преобразовать кПа в мм рт. Ст., Умножьте на 7.5).
Рис. 5. Изменения в составе альвеолярного воздуха во время нормального дыхательного цикла в состоянии покоя. Шкала слева и синяя линия указывает парциальные давления углекислого газа в кПа, а шкала справа и красная линия указывает парциальные давления кислорода, также в кПа (чтобы преобразовать кПа в мм рт. Ст., Умножьте на 7.5).  Рис. 6. Схематическое гистологическое сечение части легочной ткани, показывающее нормально вздутую альвеолу (в конце нормального выдоха) и ее стенки, также альвеолярные капилляры (показано в разрезе). Это показывает, как кровь альвеолярных капилляров полностью окружена альвеолярным воздухом. В нормальном легком человека все альвеолы вместе содержат около 3 литров альвеолярного воздуха. Все альвеолярные капилляры содержат около 100 мл крови.
Рис. 6. Схематическое гистологическое сечение части легочной ткани, показывающее нормально вздутую альвеолу (в конце нормального выдоха) и ее стенки, также альвеолярные капилляры (показано в разрезе). Это показывает, как кровь альвеолярных капилляров полностью окружена альвеолярным воздухом. В нормальном легком человека все альвеолы вместе содержат около 3 литров альвеолярного воздуха. Все альвеолярные капилляры содержат около 100 мл крови. Воздух доставляется в альвеолы небольшими дозами (так называемый дыхательный объем ) посредством дыхания в (вдох ) и выдох (выдох ) через дыхательные пути, набор относительно узких и умеренно длинных трубок, которые начинаются у носа или рта и заканчиваются в альвеолах легкие в груди. Воздух входит и выходит через один и тот же набор трубок, в которых идет в одном направлении, во время вдоха и в противоположном направлении во время выдоха.
Во время вдоха в состоянии покоя через нос поступает около 500 мл свежего воздуха. Его согревают и увлажняют, когда он течет через нос и глотку. К тому времени, когда воздух достигает трахеи, температура вдыхаемого воздуха составляет 37 ° C, и он насыщается водяным паром. По прибытии в альвеолы он разбавляется и тщательно смешивается примерно 2,5–3,0 литрами воздуха, оставшимся в альвеолах после последнего выдоха. Этот относительно большой объем воздуха, который полупостоянно присутствует в альвеолах на протяжении всего цикла дыхания, известен как функциональная остаточная емкость (FRC).
В начале вдоха дыхательные пути закрыты. наполнен наполненным альвеолярным воздухом, оставшимся после последнего выдоха. Это объем мертвого пространства, который обычно составляет около 150 мл. Это первый воздух, который повторно попадает в альвеолы при вдохе. Только после того, как воздух мертвого пространства вернулся в альвеолы, остаток дыхательного объема (500 мл - 150 мл = 350 мл) попадает в альвеолы. Поступление такого небольшого объема свежего воздуха при каждом вдохе гарантирует, что состав FRC практически не изменится во время цикла дыхания (рис. 5). Альвеолярное парциальное давление кислорода остается очень близким к 13–14 кПа (100 мм рт. Ст.), А парциальное давление углекислого газа колеблется минимально около 5,3 кПа (40 мм рт. Ст.) На протяжении дыхательного (вдоха и выдоха). Соответствующие парциальные давления кислорода и углекислого газа в окружающем (сухом) воздухе на уровне моря составляют 21 кПа (160 мм рт. Ст.) И 0,04 кПа (0,3 мм рт. Ст.) Соответственно.
 Рис. 7. Очень схематическая иллюстрация процесса газообмена легких млекопитающих, выделяющая различия между газовым составом окружающего воздуха, альвеолярным воздухом (светло-голубой), с которым уравновешивается альвеолярная капиллярная кровь, и кровью. газовое давление в легочной артерии (синяя кровь поступает в легкое слева) и венозной крови (красная кровь выходит из легкого справа). Все газовые напряжения указаны в кПа. Чтобы преобразовать его в мм рт. Ст., Умножьте на 7,5.
Рис. 7. Очень схематическая иллюстрация процесса газообмена легких млекопитающих, выделяющая различия между газовым составом окружающего воздуха, альвеолярным воздухом (светло-голубой), с которым уравновешивается альвеолярная капиллярная кровь, и кровью. газовое давление в легочной артерии (синяя кровь поступает в легкое слева) и венозной крови (красная кровь выходит из легкого справа). Все газовые напряжения указаны в кПа. Чтобы преобразовать его в мм рт. Ст., Умножьте на 7,5. Этот альвеолярный воздух, составляющий FRC, полностью окружает кровь в альвеолярных капиллярах (рис. 6). У млекопитающих газообмен происходит между этим альвеолярным воздухом (который значительно отличается от свежего воздуха) и кровью в альвеолярных капиллярах. Газы по обе стороны от газообменной мембраны уравновешиваются простым диффузией. Это гарантирует, что парциальное давление кислорода и углекислого газа в крови, покидающей альвеолярные капилляры и в соответствии с указанием циркулирующей по всему телу, будет таким же, как и в FRC.
Заметная разница между составом альвеол воздух и окружающий воздух могут поддерживаться, поскольку функциональная остаточная емкость содержится в тупиковых мешочках, соединенных с наружным воздухом длинными узкими трубками (дыхательные пути: нос, глотка, гортань, трахея, бронхи и их ветви и части до бронхиол ). Эта анатомия, а также тот факт, что легкие не опорожняются и не наполняются заново с каждым вдохом, обеспечивает млекопитающим «переносимую атмосферу», состав которой значительно отличается от современного окружающего воздуха.
Состав Воздух в FRC тщательно контролируется путем измерения парциального давления кислорода и углекислого газа в артериальной крови. Если давление газа отклоняется от нормы, возникают рефлексы, которые изменяют частоту и глубину дыхания таким образом, что нормальное состояние восстанавливается в течение секунд или минут.
Вся кровь возвращается из тканей тела в правую сторону сердца проходит через альвеолярные капилляры, прежде чем снова прокачиваться по всему телу. Проходя через легкие, кровь вступает в тесный контакт с альвеолярным воздухом, отделенным от него очень тонкой диффузионной мембраной, которая в среднем составляет всего около 2 мкм. Таким образом, давление газа в крови будет быстро уравновешиваться с давлением в альвеолах, гарантируя, что артериальная кровь, которая циркулирует во всех тканях по всему телу, имеет напряжение кислорода, равное 13−14. кПа (100 мм рт. ст.) и давление диоксида углерода 5,3 кПа (40 мм рт. ст.). Эти артериальные парциальные давления кислорода и углекислого газа контролируются гомеостатически. Повышение артериального 

Поскольку кровь, поступающая в альвеолярные капилляры, имеет
Только в результате точного поддержания состава 3 литров альвеолярного воздуха при вдохе некоторое количество углекислого газа выбрасывается в атмосферу, а некоторое количество кислорода поглощается из внешнего воздуха. Если за короткий период гипервентиляции было потеряно больше углекислого газа, чем обычно, дыхание будет замедлено или остановлено до тех пор, пока альвеолярный
Если эти гомеостаты нарушены, то респираторный ацидоз или респираторный алкалоз. В итоге это может быть компенсировано почечной корректировкой концентраций H и HCO 3 в плазме; но поскольку это требует времени, синдром гипервентиляции может, например, вызвать, когда возбуждение или тревога заставляют человека дышать быстро и тем самым, тем самым выбрасывая слишком много CO 2 из крови в наружный воздух, вызывая набор неприятных симптомов, которые вызывают в результате чрезмерно высокого pH внеклеточных жидкостей.
Кислород имеет очень низкую растворимость в воде и поэтому переносится в крови в слабом сочетании с гемоглобин. Кислород удерживается на гемоглобине четырьмя двухвалентным железом -содержими гемом группой на молекулу гемоглобина. Когда все гемовые группы несут по одной молекуле O 2 каждая, кровь считается «насыщенной» кислородом, и никакое дальнейшее повышение парциального давления кислорода не приводит к значительному увеличению кислорода в крови. Большая часть диоксида углерода в крови переносится в плазме в виде домашнего HCO 3. Однако преобразование растворенного CO 2 в HCO 3 (посредством добавления воды) слишком медленное для скорости, с которой кровь циркулирует через ткань, с одной стороны, и альвеолярной капилляры с другой. Следовательно, реакция катализируется карбоангидразой, ферментом внутри красных кровяных телец. Реакция может идти в любом направлении в зависимости от преобладающего парциального давления диоксида углерода. Небольшое количество диоксида углерода переносится белковой молекулой гемоглобина в виде карбамино. Общая концентрация углекислого газа (в форме бикарбоната, растворенного CO 2 и карбаминогрупп) в артериальной крови (то есть после того, как она уравновесится с альвеолярным воздухом) составляет около 26 мМ (или 58 мл. / 100 мл.) по сравнению с концентрацией кислорода в насыщенной артериальной крови около 9 мМ (или 20 мл / 100 мл крови).
 Рис. 8. Жабры тунца с нитями и ламелями
Рис. 8. Жабры тунца с нитями и ламелями Содержание растворенного кислорода в пресной воде составляет примерно 8–10 миллилитров на литр по сравнению с содержанием воздуха, которое составляет 210 миллилитров на литр. Вода в 800 раз плотнее воздуха и в 100 раз вязче. Следовательно, скорость диффузии кислорода в воздухе в 10 000 раз выше, чем в воде. Следовательно, использование мешкообразных легких для удаления кислорода из воды было бы недостаточно эффективным для поддержания жизни. Вместо использования легких газообмен происходит на поверхности сильно васкуляризованных жабр. Жабры - это специализированные органы, содержащие нитей, которые далее на пластинки. Пластинки <содержат301>капилляры, которые обеспечивают большую площадь коротких расстояний, так как их стенки очень тонкие. Жаберные тычинки находятся в системе обмена, чтобы фильтровать пищу и поддерживать чистоту жабр.
В жабрах используется система противотока, которая увеличивает эффективность поглощения кислорода (и потери отработанного газа). Кислородная вода втягивается через рот и проходит через жабры в одном направлении, тогда как кровь через ламели в противоположном направлении. Этот противоток поддерживает резкие меры по концентрации по всей длине каждого капилляра (см. Диаграмму в разделе «Взаимодействие с кровеносными системами» выше). Кислород может непрерывно диффундировать по градиенту в кровь, а углекислый газ - вниз по градиенту в воду. Деоксигенированная вода в итоге будет проходить через operculum (жаберное покрытие). Хотя противоточные системы обмена теоретически допускают почти полный перенос дыхательного газа с одной стороны теплообменника на другой, у рыб менее 80% кислорода в воде, протекающей по жабрам, обычно переносится в кровь.
У земноводных есть три органа, участвующих в газообмене: легкие, кожа и жабры, которые можно использовать по отдельности или в различных комбинациях. Относительная структура зависит от возраста, среды обитания и вида амфибии. Кожа амфибий и их личинок сильно васкуляризована, что приводит к относительно эффективному газообмену, когда кожа влажная. Личинки земноводных, такие как стадия преметаморфоза головастика у лягушек, также имеют наружные жабры. Жабры поглощаются телом во время метаморфоза, после чего легкие вступают во владение. Легкие обычно проще, чем у других наземных позвоночных, с небольшими внутренними перегородками и более крупными альвеолами; однако жабы, имеют больше времени на суше, имеют альвеолярную поверхность и более развитые легкие. Чтобы увеличить скорость газообмена за счет диффузии, земноводные процесс накопления на респираторной поверхности, используя, называемый буккальным насосом. Нижний этаж рта перемещается «качающим» образом, что можно наблюдать невооруженным глазом.
Все рептилии дышат легкими. У чешуйчатых (ящериц и змей ) вентиляция управляется осевой мускулатурой, но эта мускулатура также используется во время движения, поэтому некоторые плоскоклеточные полагаются на буккальную откачку для поддержания эффективности газообмена.
Из-за жесткости панцирей черепахи и черепахи значительное расширение и сжатие грудной клетки сложно. Черепахи и черепахи зависят от мышечных слоев, прикрепленных к их панцирям, которые окружают их легкие, наполняя и опорожняя их. Некоторые водные черепахи также могут закачивать воду в сильно васкуляризованный рот или клоаку для газообмена.
Крокодилы имеют структуру, похожую на диафрагмальных млекопитающих - но эта мышца создает однонаправленный поток воздуха через легкие, а не приливный поток: это больше похоже на поток воздуха, наблюдаемый у птиц, чем у млекопитающих. Во время вдоха диафрагмальная мышца втягивает печень назад, раздувая легкие в создаваемое пространство. Воздух поступает в легкие из бронхов во время вдоха, но во время выдоха воздух из легких через одностороннее движение газа достигается за счет аэродинамических клапанов в дыхательных путях.
 Рис. 10. Цикл вдох-выдох у птиц.
Рис. 10. Цикл вдох-выдох у птиц.  Рис. 9. Схематическое изображение перекрестно-точного газообменника дыхательных путей в легких птиц. Воздух вытесняется из воздушных мешков в одном направлении (справа налево на схеме) через парабронхи. Легочные капилляры окружают парабронхи, как показано на рисунке (кровь от нижнего края парабронха к верхнему на диаграмме). Кровь или воздух с высоким содержанием кислорода показаны красным цветом; Бедный кислородом воздух или кровь показаны различными оттенками пурпурно-синего цвета.
Рис. 9. Схематическое изображение перекрестно-точного газообменника дыхательных путей в легких птиц. Воздух вытесняется из воздушных мешков в одном направлении (справа налево на схеме) через парабронхи. Легочные капилляры окружают парабронхи, как показано на рисунке (кровь от нижнего края парабронха к верхнему на диаграмме). Кровь или воздух с высоким содержанием кислорода показаны красным цветом; Бедный кислородом воздух или кровь показаны различными оттенками пурпурно-синего цвета. У птиц легкие, но нет диафрагмы. В основном они полагаются на воздушные мешки для вентиляции. Эти воздушные мешочки не играют непосредственной роли в газообмене, но помогают перемещать воздух в одном направлении через поверхность газообмена в легких. Во время вдоха свежий воздух забирается из трахеи в задние воздушные мешки и в парабронхи, которые ведут из задних воздушных мешочков в легкое. Воздух, попадающий в легкие, присоединяется к воздуху, который уже находится в легких, и направляется вперед через газообменник в передние воздушные мешочки. Во время выдоха задние воздушные мешочки нагнетают воздух в те же парабронхи легких, двигаясь в том же направлении, что и во время вдоха, непрерывный газообмен независимо от цикла дыхания. Воздух, выходящий из легких во время выдоха, присоединяется к воздуху, выходящему из передних воздушных мешков (оба они состоят из «отработанного воздуха», прошедшего через газообменник), поступающего в трахею для выдоха (рис. 10). Селективное сужение бронхов в различных точках ветвления бронхов гарантирует, что воздух не будет течь через бронхи во время вдоха и выдоха, как это происходит у млекопитающих, а следует путями, описанными выше.
Однонаправленный поток воздуха, проходящий через парабронхи, обменивается дыхательными газами с поперечным током крови (рис. 9). Парциальное давление O 2(
В газообмене растений доминируют углекислый газ, кислород и водяной пар. CO. 2является единственным источником углерода для автотрофного роста посредством фотосинтеза, и когда растение активно фотосинтезирует на свету, оно будет поглощать углекислый газ, и потеря водяного пара и кислорода. Ночью растения дышат, и газообмен частично меняется на противоположный: водяной пар все еще теряется (но в меньшей степени), но теперь поглощается кислород и выделяется углекислый газ.
 Рис. 11. Стилизованное поперечное сечение листа растения эуфиллофита, показывающее ключевые органы растения, участвующие в газообмене.
Рис. 11. Стилизованное поперечное сечение листа растения эуфиллофита, показывающее ключевые органы растения, участвующие в газообмене. Газообмен растений происходит в основном через листья. Газы диффундируют в и из межклеточных пространств внутри листа через поры, называемые устьицами, которые обычно находятся на нижней поверхности листа. Газы попадают в фотосинтетическую ткань листа через растворение на влажной поверхности палисада и губчатых клеток мезофилла. Губчатые клетки мезофилла неплотно упакованы, что позволяет увеличить площадь поверхности и, следовательно, увеличить скорость газообмена. Поглощение углекислого газа обязательно приводит к некоторой потере водяного пара, потому что обе молекулы входят и выходят через одни и те же устьица, поэтому растения сталкиваются с дилеммой газообмена: получить достаточно CO. 2без потери слишком большого количества воды. Следовательно, потеря воды из других частей листа сводится к минимуму за счет восковой кутикулы на эпидермисе листа. Размер стомы регулируется открытием и закрытием двух ее замыкающих клеток : отечность этих клеток определяет состояние устьичного отверстия, а это само регулируется водным стрессом. Растения, демонстрирующие метаболизм крассулоидной кислоты, устойчивы к засухе ксерофиты и осуществляют почти весь свой газообмен в ночное время, потому что только ночью эти растения открывают устьица. Открытие устьиц только на ночь сводит к минимуму потери водяного пара, связанные с поглощением углекислого газа. Однако это происходит за счет медленного роста: растение должно хранить углекислый газ в форме яблочной кислоты для использования в течение дня, и оно не может хранить неограниченное количество.
 Рис. 12. Высокоточные измерения газообмена дают важную информацию о физиологии растений
Рис. 12. Высокоточные измерения газообмена дают важную информацию о физиологии растений Измерения газообмена являются важными инструментами в науке о растениях: обычно это включает герметизацию растения (или части растения) в камере и изменения концентрации углекислого газа с помощью инфракрасного газоанализатора . Если условия окружающей среды (влажность, концентрация CO. 2, свет и температура ) полностью контролируются, измерения поглощения CO. 2и выделения воды дают важную информацию о CO.. 2ассимиляция и транспирация коэффициенты. Межклеточная концентрация CO. 2дает важную информацию о фотосинтетическом состоянии растений. В определенных обстоятельствах можно использовать более простые методы: индикатор гидрокарбоната можно использовать для контроля потребления CO. 2в растворе, содержащем один лист растения, при разных уровнях интенсивности света и генерации кислорода водорослями. Элодея можно измерить, просто собрав газ в погруженную пробирку, содержащую небольшой кусочек растения.
Механизм газообмена у беспозвоночных