
Войти
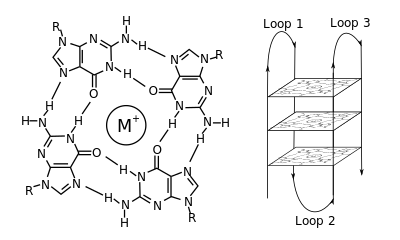 Структура G-квадруплекса. Слева: G-тетрада. Справа: внутримолекулярный комплекс G4.
Структура G-квадруплекса. Слева: G-тетрада. Справа: внутримолекулярный комплекс G4. В молекулярной биологии G-квадруплексвторичные структуры (G4) образуются в нуклеиновых кислотах с помощью последовательностей, богатых гуанином. Они имеют спиралевидную форму и содержат тетрады гуанина, которые могут образовывать одну, две или четыре нити. Мономолекулярные формы встречаются в природе вблизи концов хромосом, более известных как теломерные области, транскрипции множества генов, как у микробов, так и у позвоночных, включая онкогены у людей. Четыре гуаниновых основания могут связываться посредством водородной связи Хугстина с образованием плоской квадратной структуры, называемой тетрадой гуанина (G-тетрада или G-квартет), и две или более тетрад гуанина (от G - ракты, непрерывные прогоны гуанина) могут складываться друг на друга, образуя G-квадруплекс.
Размещение и связь с образованием G-квадрупсов не случайны и очень необычно функциональным целям. Структура квадруплекса стабилизируется за счет присутствия катиона, особенно калия, который находится в центральном канале между каждым парой тетрад. Они могут состоять из ДНК, РНК, LNA и PNA и могут быть внутримолекулярными, бимолекулярный или тетрамолекулярный. В зависимости от цепей или частей цепочки, образующих тетрады, структуры могут быть стимулы как параллельные или антипараллельные. Структуры G-квадруплексов могут быть предсказаны с помощью вычислений на основе мотивов последовательностей ДНК или РНК, но их фактические структуры могут быть сильно изменены внутри и между мотивами, которые могут быть более 100000 на геном. Их деятельность основных генетических процессов является активной областью исследований в области регуляции генов и функциональной геномики.
Идентификация структур с высокой ассоциацией гуанина стала очевидной в начале 1960-х годов, благодаря идентификации гелеобразных веществ, связанных с гуанинами. Более конкретно, это исследование детализировало четырехцепочечные структуры ДНК с высокой ассоциацией гуанинов, которые были идентифицированы в эукариотических теломерных областях ДНК в 1980-х годах. Важность открытия структуры G-квадруплекса была описана в заявлении: «Если G-квадруплексы так легко образуются in vitro, природа найдет способ их использования in vivo» - Аарон Клаг, лауреат Нобелевской программы по химии (1982). Интерес к функциям G-квадруплексов in vivo возникает после того, как крупномасштабный полногеномный анализ показал преобладание последовательностей, образующих G-квадруплекс (pG4), в промоторах генов человека, шимпанзе, мыши и крысы, представленных в Первом международном обзоре G. -квадруплекс Встреча состоялась в апреле 2007 года в Луисвилле, Кентукки. В 2006 году сообщалось о преобладании G-квадруплексов в промоторах нескольких бактериальных геномов, которые предсказывают опосредованную G-квадруплексами регуляцию генов. При большом количестве G-квадруплексов in vivo эти структуры играют биологически значимую роль благодаря взаимодействию с промоторными областями онкогенов и теломерными областями цепей ДНК. Текущие исследования заключаются в определении биологической функции этих структур G-квадруплексов для конкретных терапевтических методов лечения рака, основанных на взаимодействии с G-квадруплексами.
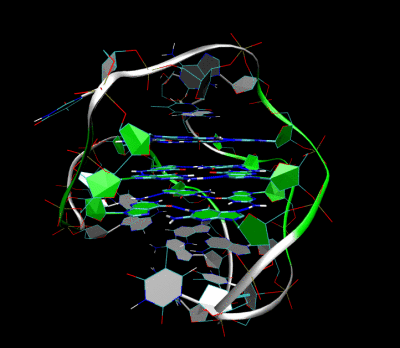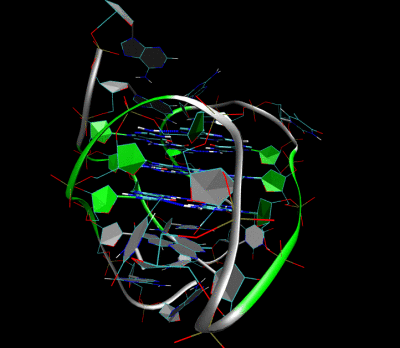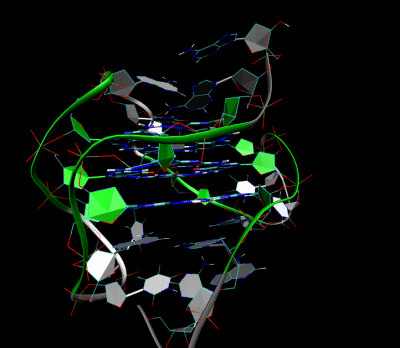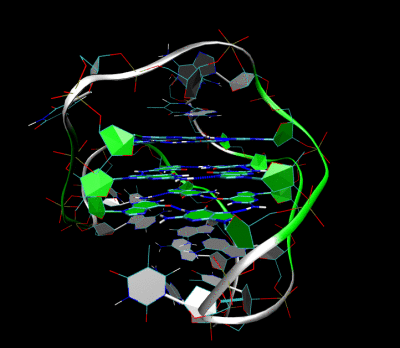 Трехмерная структура внутримолекулярного теломерного G-квадруплекса человека в растворе калия. Костяк представлен трубкой. В центре этой структуры находятся три слоя G-тетрад. Водородные связи в этих слоях представлены синими пунктирными линиями. (PDB : 2HY9 )
Трехмерная структура внутримолекулярного теломерного G-квадруплекса человека в растворе калия. Костяк представлен трубкой. В центре этой структуры находятся три слоя G-тетрад. Водородные связи в этих слоях представлены синими пунктирными линиями. (PDB : 2HY9 )Длина последовательностей нуклеиновых кислот, участвующих в образовании тетрад, определяет, как складывается квадруплекс. Короткие последовательность, состоящие только из одного непрерывного ряда трех или Такие квадруплексные структуры требуют четырех отдельных цепей для образования квадруплекса. Более длинные следы, содержащие два ряда из трех или более оснований гуанина, где области гуанина разделены одно или несколько оснований требуют только двух последовательностей для обеспечения достаточного количества оснований гуанина для квадрупрупа лекса. Эти структуры, образованные из двух отдельных G-богатых цепей, называются бимолекулярными квадруплексами. Наконец, представлены четыре расстояния. Ряды оснований гуанина сами по себе могут образовывать стабильные квадруплексные структуры, и квадруплекс, полностью сформированный из одной цепи, называется внутримолекулярным квадруплексом.
В зависимости от того, как отдельные серии оснований расположены в бимолекулярном или внутримолекулярном квадруплекс, квадруплекс может принимать одну из множества топологий с различными конфигурациями петель. Если все цепи ДНК движутся в одном направлении, квадруплекс называется параллельным. Пропеллерного типа и располагаться по бокам квадруплекса. Если один или несколько рядов оснований гуанина имеют направление 5’-3 ’, противоположное рядов оснований гуанина, считается, что квадруплекс принял антипараллельную топологию. Петли, соединяющие ряды оснований гуанина во внутримолекулярных антипараллельных квадруплексах, являются либо диагональными, соединяющими два диагонально противоположных ряда оснований гуанина, либо петлями латерального (на ребро) типа, соединяющими два соседних ряда пар оснований гуанина.
В квадруплексах, образованных из двухцепочечной ДНК, также обсуждались возможные межцепочечные топологии. Межцепочечные квадруплексы содержат гуанины, происходящие из цепей дцДНК.
секвенирования генома человека было обнаружено множество богатых гуаниномных последовательностей, которые могли образовывать квадруплексы. В клетки типа клетки и клеточного цикла, опосредующие факторы, такие как ДНК-связывающие белки на зависимости, состоящие из ДНК, плотно намотанной вокруг белков гистона, а также другие условия окружающей среды и стрессы на динамическое формирование квадрупсов. Например, количественные оценки термодинамики молекулярного краудинга показывают, что антипараллельный g-квадруплекс стабилизируется молекулярным краудингом. Этот эффект, по-видимому, опосредован изменением гидратации ДНК и его привязки пары оснований Хугстина. Эти квадруплексы, по-видимому, легко находятся на концах хромосомы. Кроме того, склонность к образованию g-квадруплексов во время транскрипции в последовательности РНК, способных образовывать взаимоисключающие структуры шпильки или G-квадруплекса, сильно зависит от положения последовательности, образующей шпильку.
Ферменты репарации естественным образом распознают концы линейных хромосом как поврежденную ДНК и обрабатывают их как таковые, оказывая вредное воздействие на клетку, необходимы четкая передача сигналов и жесткая регуляция на концах линейных хромосом. Теломеры обеспечивают эту сигнализацию. Теломеры, защищенные гуанином и склонные к образованию g-квадруплексов, безопасны на концевых концах хромосом и обеспечивают целостность генома, защищая эти уязвимые концевые концевые концы от нестабильности.
Эти теломерные области характеризуются постоянными участками двухцепочечных повторов CCCTAA: TTAGGG. Повторы заканчиваются 3’-выступом из 10-50 однонитевых повторов ТТАГГГ. Гетеродимерный комплексный рибонуклеопротеиновый фермент теломераза сэр повторы TTAGGG на 3’-конце цепей ДНК. На этих 3’-концевых выступах может образоваться вторичная структура, например G-квадруплексы, если выступать длиннее четырех повторов TTAGGG. Присутствие этих предотвращает удлинение теломер за счет теломеразного комплекса.
Теломерные повторы у различных организмов, как было показано, образуют эти квадруплексные структуры in vitro, и показано, что они образуют in vivo. Теломерный повтор человека (который является одинаковым для всех позвоночных ) состоит из множества повторов секвенированного (GGTTAG), и образованные этой структурой, могут иметь форму бусинок размером от 5 до 8 нм. по размеру и были хорошо изучены с помощью ЯМР, ПЭМ и X-рентгеновского определения кристаллической структуры. Было показано, что эти квадлексов в теломерах снижает активность фермента теломеразы образование, которое отвечает за поддержание прочности теломер и участвует примерно в 85% всех раки. Это активная цель открытия лекарств, в том числе теломестатин.
Квадруплексы присутствуют не в теломере и в других местах. Анализ геномов человека, шимпанзе, мыши и крысы показал огромное количество образующих G-квадруплекс (pG4) последовательностей в нетеломерных областях. Большое количество нетеломерных G-квадруплексов было обнаружено в промоторах генов и было законсервировано для всех видов. Точно так же большое количество G-квадруплексов было обнаружено в E. coli и сотнях других микробных геномов. Здесь, как и у позвоночных, G-квадруплексы были также обогащены промоторами генов. Хотя эти исследования предсказали опосредованную G-квадруплексом регуляцию генов, маловероятно, что все pG4 будут образовываться in vivo. протоонкоген c-myc образует квадруплекс в гиперчувствительной области нуклеазы, критической для активности гена. Другие гены, образующие G-квадруплексы в их промоторных включают области, куриный β-глобин ген, человеческий убиквитин -лигаза RFP2 и протоонкогены c-kit, bcl-2, VEGF, H- ras и N-ras.
Геном были проведены исследования на основе правил складывания квадруплексов, которые выявили 376000 предполагаемых квадруплексных последовательностей (PQS) в человеке геном, хотя не все из них, вероятно, образуются in vivo. Аналогичные исследования выявили предполагаемые G-квадруплексы у прокариот. Существует несколько моделей того, как квадлексы могут влиять на активность генов посредством повышения регуляции, либо понижающей регуляции. Одна модель ниже, с образованием G-квадруплекса в промоторе или рядом с, блокирующим транскрипцию гена и, следовательно, деактивирующим его. В другой модели квадруплекс, образованный на некодирующей цепи ДНК, помогает поддерживать открытую конформацию кодирующей цепи ДНК и усиливать экспрессию соответствующей гена.
Было высказано предположение, что образование квадруплекса играет роль в переключении тяжелой цепи иммуноглобулина. По мере развития клеток развивались механизмы разрешения (т. Е. Раскручивания) образующихся квадруплексов. Образование квадруплексов может быть опасным для клетки; геликазы WRN и белок синдрома Блума обладает высоким сродством к разрешению G-квадруплексов ДНК. Геликаза DEAH / RHA, DHX36, также была идентифицирована как ключевая резольваза G-квадруплекса. В 2009 году было обнаружено, что белок-супрессор метастазов NM23H2 (также известный как NME2) напрямую взаимодействует с G-квадруплексом в промоторе гена c-myc и транскрипционно регулирует c-myc. Совсем недавно было сообщено, что NM23H2 взаимодействует с G-квадруплексом в промоторе гена теломеразы человека (hTERT) и регулирует экспрессию hTERT. В 2019 году было показано, что теломер-связывающий фактор-2 (TRF2 или TERF2) связывается с тысячами нетеломерные G-квадруплексы в геноме человека TRF2 ChIP-seq. Существует множество исследований, в которых квадрупсы участвуют как в позитивной, так и в негативной регуляции транскрипции, включая эпигенетическую регуляцию генов, подобных hTERT. Сообщалось также, что функция G-квадруплексов позволяет запрограммировать рекомбинацию тяжелого генов иммунологлобина и системы вариаций антигенного пилина патогоглобина Neisseria. Роли квадруплексной структуры в управлении трансляцией изучены недостаточно. Прямая визуализация структур G-квадруплексов в клетках человека, а также сокристаллическая структура РНК-геликазы, структура с G-квадруплексом, предоставила важные подтверждения их значимости для клеточной биологии. Возможные положительные и отрицательные роли квадруплексов в репликации и функции теломер остаются спорными. Т-петли и G-квадлексы описываются как две третичные структуры ДНК, которые защищают концы теломер и регулируют длину теломер.
Последовательности, образующие G-квадруплекс, представляют собой преобладает в эукариотических клетках, особенно в теломерах, 5` нетранслируемых цепях и горячих точках транслокации. G-легко разворачиваются с помощью геликазы. Однако в раковых клетках, которые имеют мутировавшую геликазу, эти комплексы не могут быть размотаны и потенциальному потенциальному повреждению клетки. Это вызывает репликацию поврежденных и раковых клеток. Для терапевтических достижений стабилизация G-квадруплексов раковых клеток может подавлять рост и репликацию клеток, что приводит к гибели клеток.
Наряду с ассоциацией G-квадруплексов в теломерных области ДНК, G-квадруплексные структуры были идентифицированы в различных промоторных областях человеческого прото- онкогена. Соответствующие структуры G-квадруплекса с параллельными цепями ДНК, как правило, соответствуют структурам G-квадруплекса с параллельными цепями ДНК. Некоторые из этих онкогенов включают c-KIT, PDGF-A, c-Myc и VEGF, что свидетельствует о важности этой вторичной структуры для роста и развития рака. Несмотря на то, что структура структуры G-квадруплекса до некоторой области для разных промоторных областей онкогенов, последовательная стабилизация этих структур была обнаружена при развитии рака. Текущие терапевты активно фокусируются на стабилизации структур G-квадруплекса для остановки нерегулируемого роста и деления клеток.
Одна конкретная область гена, путь c-myc, играет важную роль в регуляции языкового продукта c-Myc. С этим продуктом белка c-Myc работает процесс апоптоза и роста или развития клеток, а также в качестве транскрипционного контроля обратного транскриптазы теломеразы человека. Было показано, что взаимодействие G-квадруплекса промотора c-Myc с NM23H2 регулирует c-Myc в раковых клетках в 2009 г.
Регуляция c-myc через обратную транскриптазу теломеразы человека (hTERT) также напрямую регулируется через промотор G-квадруплекс путем взаимодействия с фактором транскрипции NM23H2, где эпигенетические модификации зависели от ассоциации NM23H2-G-квадруплекс. Недавно сообщалось, что эпигенетическая регуляция hTERT опосредована взаимодействием G-квадруплекса промотора hTERT с теломерным фактором TRF2.
Другой путь гена связан с геномом VEGF, фактором роста эндотелия сосудов, который по-прежнему участвует в процессе ангиогенез или образование новых кровеносных сосудов. Формирование внутримолекулярной структуры G-квадруплекса было показано посредством исследований полипуринового тракта промоторной области гена VEGF. Благодаря недавним исследованиям роли функции G-квадруплекса in vivo показано, что стабилизация структур G-квадруплекса регулирует транскрипцию гена VEGF с ингибированием факторов транскрипции в этом пути. Внутримолекулярные структуры G-квадруплексов образуются в основном за счет обильной гуаниновой последовательности в промоторной области этого специфического пути. Ген CDKN1A (также известный как p21), ген ингибитора-1 киназы контрольной точки циклинзависимого клеточного цикла, содержит промоторный G-квадруплекс. Взаимодействие этого G-квадруплекса с TRF2 (также известным как TERF2) привело к эпигенетической регуляции p21, что было протестировано с использованием лиганда, связывающего G-квадруплекс 360A.
Фактор, индуцируемый гипоксией 1ɑ, HIF-1ɑ, остается участвует в передаче сигналов рака через его связывание с элементом ответа на гипоксию, HRE, в присутствии гипоксии, чтобы начать процесс ангиогенеза. Благодаря недавним исследованиям этого специфического генного пути участок полипурина и полипиримидина позволяет транскрипцию этого специфического гена и формирование внутримолекулярной структуры G-квадруплекса. Однако необходимы дополнительные исследования, чтобы определить, регулирует ли образование G-квадруплекса экспрессию этого гена положительным или отрицательным образом.
Онкоген c-kit имеет дело с путем, который кодирует RTK, который был Показано, что при некоторых типах рака уровень экспрессии повышен. Богатая гуаниновая последовательность этой промоторной области показала способность образовывать множество квадруплексов. Текущие исследования этого пути сосредоточены на обнаружении биологической функции этого специфического образования квадруплекса на пути c-kit, в то время как эта квадруплексная последовательность была замечена у различных видов.
Онкоген RET участвует в транскрипции киназа, которая широко используется при некоторых типах рака. Богатая гуанином последовательность в промоторной области этого пути обусловливает необходимость базовой транскрипции этой рецепторной тирозинкиназы. При определенных типах рака уровень экспрессии белка RET повышен. Исследование этого пути показало образование G-квадруплекса в промоторной области и подходящую мишень для терапевтического лечения.
Другой путь онкогена с участием PDGF-A, фактора роста тромбоцитов, включает процесс раны исцеление и функционирование как митогенные факторы роста клеток. Высокие уровни экспрессии PDGF были связаны с повышенным ростом клеток и раком. Присутствие богатой гуанином последовательности в промоторной области PDGF-A продемонстрировало способность образовывать внутримолекулярные параллельные G-квадруплексные структуры и, как предполагается, играет роль в регуляции транскрипции PDGF-A. Однако исследования также выявили наличие структур G-квадруплексов в этой области из-за взаимодействия TMPyP4 с этой промоторной последовательностью.
Теломеры обычно состоят из G-квадруплексов и остаются важными объектами терапевтических исследований и открытий. Эти комплексы обладают высоким сродством к порфириновым кольцам, что делает их эффективными противораковыми агентами. Однако использование TMPyP4 ограничено из-за его неизбирательности по отношению к теломерам раковых клеток и нормальной двухцепочечной ДНК (дцДНК). Чтобы решить эту проблему, был синтезирован аналог TMPyP4, известный как 5Me, который нацелен только на G-квадруплексную ДНК, которая ингибирует рост рака более эффективно, чем TMPyP4.
Дизайн и разработка лигандов остается важной областью исследований терапевтических реагентов из-за обилие G-квадруплексов и их множественные конформационные различия. Один тип лиганда, включающий производное хиндолина, SYUIQ-05, использует стабилизацию G-квадруплексов в промоторных областях для ингибирования продукции как белкового продукта c-Myc, так и обратной транскриптазытеломеразы человека (hTERT). Этот путь нацеливания на эту область приводит к отсутствию элонгации теломеразы, что приводит к остановке развития клеток. По-прежнему необходимы дальнейшие исследования для открытия единственного гена-мишени для минимизации нежелательной реактивности с более эффективной противоопухолевой активностью.
Один из способов индуцирования или стабилизации образования G-квадруплекса - это: молекула, которая может связываться со структурой G-квадруплекса. Ряд лигандов, которые могут быть как небольшие молекулами, так и белками, связываются с G-квадруплексом. Эти лиганды могут быть встречающимися в природе или синтетическими. Это становится все более обширной областью исследований в области генетики, биохимии и фармакологии.
Катионные порфирины, как было показано, интеркалятивно связываются с G-квадруплексами, а также с молекулами теломестатин.
Связывание лигандов с G-квадруплексами жизненно важно для анти- рака, потому что G-квадруплексы обычно находятся в горячих точках транслокаций. MM41, лиганд, который селективно связывается с квадруплексом на промоторе BCL-2, имеет форму центрального ядра и 4 боковых цепей, пространственно разветвляющихся. Форма лиганда жизненно важна, потому что она близко соответствует квадруплексу, который состоит из квартетов, и петлям из нуклеиновых кислот, удерживающих его вместе. При связывании хромофор MM41 центральный кварту 3’-концевого G-квартета, а боковые цепи лиганда связаны с элементами квадруплекса. Квартет и хромофор связаны π-π связью, в то время как боковые цепи и петли не связаны, находятся в непосредственной близости. Что делает это связывание прочным, так это текучесть в положении петель для лучшего связывания с боковыми цепями лиганда.
TMPyP4, катионный порфирин, более известный G4-связывающим лигандом, помогает подавлять c-Myc. Способ, TMPyP4 связывается с G4, аналогичным MM41, с кольцом, укладывающимся в стек на внешний G-квартет, а боковые цепи связаны с петлями G4.
При разработке лигандов для связывания с G-квадруплексами, лиганды обладают более высоким сродством к параллельным G-квадруплексам. Было обнаружено, что лиганды с меньшими боковыми цепями лучше связываются с квадратичными цепями, потому что лиганды меньшего размера имеют более концентрированную электронную плотность. Кроме того, водородные связи лигандов с меньшими боковыми цепями короче и, следовательно, прочнее. Лиганды с подвижными боковыми цепями, которые могут вращаться вокруг своего центрального хромофора, сильнее связаны с G-квадлексами, поскольку конформация петель G4 и боковых цепей лигандов может выравниваться.
Выявление и прогнозирование последовательностей, способных образовывать квадруплексы, являются важными инструментами для понимания их роли. Как правило, для поиска используется внутрицепочечных образующих квадруплекс последовательностей используется простое сопоставление с образцом: d (G 3+N1-7 G3+N1-7 G3+N1-7 G3+), где N - любое нуклеотидное основание (включая гуанин ). Это правило широко используется в онлайн-алгоритмах . Хотя это правило идентифицирует сайты образования G-квадруплекса, оно также идентифицирует подмножество несовершенных гомопуриновых зеркальных повторов, способных к образованиюплекса и образования i-мотивации С-цепи. Более того, эти системы также обладают способностью образовывать скользящие и свернутые структуры, которые являются неявными промежуточными продуктами в образовании как квадруплексных, так и триплексных структур ДНК. В одном исследовании было обнаружено, что наблюдаемое количество на пару оснований (то есть частота) эти мотивы быстро увеличились в эуметазоа, для которого доступны полные геномные модели. Это говорит о том, что может находиться под положительным отбором, возможно благодаря эволюции систем, способных подавлять образование не-B-структур.
Был разработан ряд экспериментальных методов для поддержки вычислительного предсказания G-квадруплексов. Эти методы можно в широком смысле разделить на два класса: биофизические и биохимические.
Биохимические методы были использованы для исследования образования G-квадруплекса в контексте более длинной последовательности. В анализе остановки ДНК-полимеразы образование G-квадруплекса в матрице ДНК может действовать как препятствие и остановку полимеразы, что останавливает удлинение праймера. Диметилсульфат (DMS) с последующим анализом расщепления пиперидина основан на том факте, что образование G-квадруплекса будет запрещать метилирование гуанина N7, вызванное DMS, что приводит к паттерну защиты, наблюдаемому в области G-квадруплекса ДНК после пиперидина.
Топология структуры G-квадруплекса определена путем мониторинга сигналов положительного или отрицательного кругового дихроизма (КД) на определенных длинах волн. Параллельные G-квадруплексы имеют отрицательные и положительные сигналы CD на 240 и 262 нм, соответственно, тогда как антипараллельные G-квадруплексы размещают эти сигналы на 262 и 295 нм соответственно. Чтобы проверить образование G-квадруплекса, необходимо также провести эксперименты с CD в условиях стабилизации не-G-квадруплекса (Li +) и стабилизации G-квадруплекса (например, K + или с лигандами G-квадруплекса) и сканировать в сторону дальней УФ-области. (180–230 нм). Точно так же термостабильность структуры G-квадруплекса можно определить, наблюдая УФ-сигнал при 295 нм. При использовании G-квадруплекса УФ-поглощение при 295 нм уменьшается, что приводит к гипохромному сдвигу, который является отличительной характеристикой структуры G-квадруплекса. Другой подход к обнаружению G-квадруплексов включает методы на основе нанопор. Во-первых, было показано, что можно выявить G-квадруплексы на основе исключения размера и специфического взаимодействия G-квадруплекса и белковой нанополости. Новый подход сочетает в себе ДНК-нанотехнологию для безметочного обнаружения G-квадруплексов, их картирования на дцДНК и для мониторинга образования G-квадруплексов.
G-квадруплексы вовлечены в неврологические расстройства посредством двух основных механизмов. Первый заключается в увеличении G-повторов внутри генов, которые вызывают образование структур G-квадлексов, которые непосредственно вызывают заболевание, как в случае с геном C9orf72 и боковым амиотрофическим склерозом (БАС) или лобно-височная деменция (FTD). Второй механизм связан с мутациями, которые на экспрессию связывающих G-квадлекс белков, как видно на примере гена умственной отсталости 1 (FMR1) и синдрома ломкой X.
гена C9orf72 кодирует белок C9orf72, который обнаруживается по всему мозгу в нейрональной цитоплазме и на пресинаптических окончаниях. Мутации гена C9orf72 связаны с развитием FTD и ALS. Эти два заболевания вызывают связь с повторами GGGGCC (G 4C2) в 1-м интроне гена C9orf72. Нормальные люди обычно имеют от 2 до 8 повторов G 4C2, но люди с FTD или ALS от 500 до нескольких тысяч повторов G 4C2. Было показано, что транскрибируемая РНК этих повторов формирует стабильные G-квадлексы, причем доказательства показывают, что повторы G 4C2в ДНК также обладают способностью образовывать смешанные параллельные-антипараллельные G-квадруплексные структуры. Было показано, что эти РНК-транскрипты, содержащие повторы G 4C2, связывают и разделяют широкий спектр спектра, включая нуклеолин. Нуклеолин участвует в синтезе и созревании рибосом в ядре, а разделенными нарушающими нуклеолина мутировавшими РНК-транскриптами вызывает ядрышковую функцию и синтез рибосомной РНК.
белок умственной отсталости Fragile X (FMRP)) представляет собой широко экспрессируемый белок, кодируемый геном FMR1, который связывается со вторичными структурами G-квадруплекса в нейронах и участвует в синаптической пластичности. FMRP действует как негативный регулятор трансляции, и его связывание стабилизирует структуру G-квадруплекса в транскриптах мРНК, ингибируя удлинение мРНК рибосомы в дендрите нейрона и контролируемое время экспрессии транскрипта.. Мутации этого гена могут вызвать развитие синдрома ломкой Х-хромосомы, аутизма и других неврологических расстройств. В частности, синдром ломкой Х-хромосомы вызван от 50 до более 200 повторов CGG в экзоне 13 гена FMR1. Эта экспансия повторов обеспечивает метилированию ДНК и другим эпигенетическим гетерохроматиновым модификациям FMR1, которые предотвращают транскрипцию гена, что приводит к патологически низкому уровню FMRP.
Антисмысловые вмешательства и низкомолекулярные лиганды являются общими стратегиями, используемыми для воздействия на неврологические заболевания, связанные с повторами экспансии G-квадруплекса. Следовательно, эти методы особенно полезны для лечения неврологических заболеваний, которые имеют механизм усиления функции, когда есть измененный генный продукт имеет новую функцию или новую экспрессию гена; это было обнаружено в C9orf72 (открытая рамка считывания 72 хромосомы 9).
Антисмысловая терапия - это процесс, с помощью которого синтезированные цепи нуклеиновых кислот используются для и специфического связывания с мРНК вырабатывается определенным геном, который его инактивирует. Антисмысловые олигонуклеотиды (ASO) обычно используются для нацеливания на РНК C9orf72 в области повторения экспансии G-квадруплекса GGGGCC, что снижает токсичность в клеточных моделях C9orf72. ASO ранее использовались для восстановления нормальных фенотипов при других неврологических заболеваниях, имели механизмы усиления функций, с той лишь разницей, что они использовались в отсутствие областей повторов расширения G-квадруплекса.
Еще один широко Используемый метод заключается в использовании низкомолекулярных лигандов . Их можно использовать для нацеливания в области G-квадруплекса, вызывающие неврологические расстройства. Существует 1000 различных G-квадруплексных лигандов, которые можно использовать через свои ароматические кольца ; это позволяет низкомолекулярным лигандам укладываться на плоские концевые тетрады в области G-квадруплекса. Недостатком использования в качестве терапевтического метода низкочастотной специфичности G-квадруплексов в их первичных последовательностях, ориентации, термодинамической стабильности и стехиометрии цепи нуклеиновой кислоты. На данный момент ни один низкомолекулярный лиганд не может быть полностью специфичным для одной G-квадруплекса. Однако катионный порфирин, известный как TMPyP4, способен связываться с повторяющейся областью C9orf72 GGGGCC, что заставляет область повторения G-квадруплекса разворачиваться и терять свои взаимодействия с белками, что приводит к потере функциональности. Низкомолекулярные лиганды, состоящие в основном из свинца, также могут быть нацелены на области повторов GGGGCC и в конечном итоге снижают как связанную с повторами трансляцию не-ATG, так и фокусы РНК в нейронных клетках, полученных от пациентов с боковым амиотрофическим склерозом (БАС). Это свидетельствует о том, что низкомолекулярные лиганды представляют собой эффективный и действенный процесс нацеливания на области GGGGCC, и что специфичность связывания низкомолекулярных лигандов является достижимой целью для научного сообщества.
Комплексы металлов обладают рядом свойств, которые делают их особенно подходящими в качестве связывающих ДНК G4 и, следовательно, в качестве потенциальных лекарств. Хотя металл играет в значительной степени структурную роль в большинстве связывающих G4, есть также примеры, когда он непосредственно взаимодействует с G4 посредством электростатических взаимодействий или прямой координации с азотистыми основаниями.