
Войти
 Филогенетическое дерево, основанное на генах rRNA , показывающий три жизненных домена : бактерии, археи и эукариоты. Черная ветвь внизу филогенетического древа соединяет три ветви живых организмов с последним универсальным общим предком. В отсутствие внешней группы корень является спекулятивным.
Филогенетическое дерево, основанное на генах rRNA , показывающий три жизненных домена : бактерии, археи и эукариоты. Черная ветвь внизу филогенетического древа соединяет три ветви живых организмов с последним универсальным общим предком. В отсутствие внешней группы корень является спекулятивным.  Автоматически созданное древо жизни с высоким разрешением, основанное на полностью секвенированных геномах.
Автоматически созданное древо жизни с высоким разрешением, основанное на полностью секвенированных геномах. A филогенетическое дерево или эволюционное дерево - это ветвящаяся диаграмма или «дерево », показывающая эволюционные отношения между различными биологическими видами или другими объектами - их филогения () - на основе сходства и различий в их физических или генетических характеристиках. Вся жизнь на Земле является частью единого филогенетического дерева, что указывает на общее происхождение.
В корневом филогенетическом дереве каждый узел с потомками представляет предполагаемого последнего общего предка этих потомков, а длины ребер в некоторых деревьях можно интерпретировать как оценки времени. Каждый узел называется таксономической единицей. Внутренние узлы обычно называют гипотетическими таксономическими единицами, поскольку их нельзя наблюдать напрямую. Деревья полезны в таких областях биологии, как биоинформатика, систематика и филогенетика. Некорневые деревья иллюстрируют только родство листовых узлов и не требуют, чтобы предковый корень был известен или предполагался.
Идея «древа жизни » возникла из древних представлений о лестничном переходе от низших к высшим формам жизни (например, в Великой Цепи Бытия ). Ранние изображения «ветвящихся» филогенетических деревьев включают «палеонтологическую карту», показывающую геологические взаимоотношения между растениями и животными в книге «Элементарная геология» Эдварда Хичкока (первое издание: 1840 г.).
Чарльз Дарвин (1859) также произвел одну из первых иллюстраций и чрезвычайно популяризировал понятие эволюционного «дерева» в своей основополагающей книге Происхождение видов. Спустя столетие биологи-эволюционисты до сих пор используют древовидные диаграммы для изображения эволюции, потому что такие диаграммы эффективно передают идею, что видообразование происходит через адаптивное и полу случайное разбиение родословных. Со временем классификация видов стала менее статичной и более динамичной.
Термин филогенетика, или филогения, происходит от двух древнегреческих слов φῦλον (phûlon), означающих «раса, происхождение» и γένεσις (génesis), что означает «происхождение, источник».
 Филогенетическое дерево с корнями, оптимизированное для слепых. Самая низкая точка дерева - корень, который символизирует всеобщего общего предка всех живых существ. Дерево разветвляется на три основные группы: бактерии (левая ветвь, буквы от a до i), археи (средняя ветвь, буквы от j до p) и эукариоты (правая ветвь, буквы от q до z). Каждая буква соответствует группе организмов, перечисленных под этим описанием. Эти буквы и описание следует преобразовать в шрифт Брайля и распечатать на принтере Брайля. Фигуру можно напечатать на 3D-принтере, скопировав файл png и используя Cura или другое программное обеспечение для генерации G-кода для 3D-печати.
Филогенетическое дерево с корнями, оптимизированное для слепых. Самая низкая точка дерева - корень, который символизирует всеобщего общего предка всех живых существ. Дерево разветвляется на три основные группы: бактерии (левая ветвь, буквы от a до i), археи (средняя ветвь, буквы от j до p) и эукариоты (правая ветвь, буквы от q до z). Каждая буква соответствует группе организмов, перечисленных под этим описанием. Эти буквы и описание следует преобразовать в шрифт Брайля и распечатать на принтере Брайля. Фигуру можно напечатать на 3D-принтере, скопировав файл png и используя Cura или другое программное обеспечение для генерации G-кода для 3D-печати. Филогенетическое дерево с корнями (см. Два рисунка вверху) - это направленный дерево с уникальным узлом - корнем - соответствующим (обычно условно исчисленным ) самому последнему общему предку всех объектов в листьях дерева. Корневой узел не имеет родительского узла, но служит родительским для всех остальных узлов в дереве. Таким образом, корень является узлом степени 2, тогда как другие внутренние узлы имеют минимальную степень 3 (где «степень» здесь относится к общему количеству входящих и исходящих ребер).
Наиболее распространенный метод для укоренения деревьев является использование спорного внешней группы -близких достаточно, чтобы позволить вывод из данных признака или молекулярной последовательности, но достаточно далеко, чтобы быть ясно внешней группой.
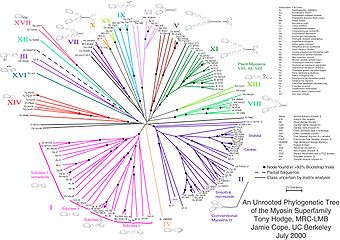 Некорневое филогенетическое дерево для миозина, суперсемейство белков.
Некорневое филогенетическое дерево для миозина, суперсемейство белков.Некорневые деревья иллюстрируют родство листовых узлов без делать предположения о происхождении. Они не требуют, чтобы предковый корень был известен или предполагался. Некорневые деревья всегда можно сгенерировать из корневых, просто опуская корень. Напротив, для определения корня дерева без корней требуются некоторые средства определения происхождения. Обычно это делается путем включения внешней группы во входные данные, чтобы корень обязательно находился между внешней группой и остальными таксонами в дереве, или путем введения дополнительных предположений об относительной скорости эволюции в каждой ветви, например, в приложении. гипотезы молекулярных часов .
И корневые, и некорневые деревья могут быть раздвоенными или многократными. У корневого бифуркационного дерева есть ровно два потомка, возникающих из каждого внутреннего узла (то есть, оно формирует двоичное дерево ), а бифуркационное дерево без корня принимает форму некорневого двоичное дерево, свободное дерево с ровно тремя соседями в каждом внутреннем узле. Напротив, у корневого мультифуркационного дерева может быть более двух дочерних элементов на некоторых узлах, а у некорневого мультифуркационного дерева может быть более трех соседей на некоторых узлах.
И корневые, и некорневые деревья могут быть помечены или немаркированы. Помеченное дерево имеет определенные значения, присвоенные его листьям, в то время как немаркированное дерево, иногда называемое формой дерева, определяет только топологию. Некоторые деревья на основе последовательностей, построенные из небольшого геномного локуса, такие как Phylotree, имеют внутренние узлы, помеченные предполагаемыми предковыми гаплотипами.
 Увеличение общего количества филогенетических деревьев в зависимости от количества помеченных листьев: бинарные деревья без корней (синие ромбы), бинарные деревья с корнями (красные кружки) и многоцветные или бинарные деревья с корнями деревья (зеленый цвет: треугольники). Масштаб по оси Y равен логарифмическому.
Увеличение общего количества филогенетических деревьев в зависимости от количества помеченных листьев: бинарные деревья без корней (синие ромбы), бинарные деревья с корнями (красные кружки) и многоцветные или бинарные деревья с корнями деревья (зеленый цвет: треугольники). Масштаб по оси Y равен логарифмическому.. Количество возможных деревьев для заданного количества листовых узлов зависит от конкретного типа дерева, но всегда имеется больше помеченных деревьев, чем немаркированных, более разнообразных, чем раздвоенных, и более укорененные, чем некорневые деревья. Последнее различие является наиболее биологически значимым; он возникает потому, что на неукорененном дереве есть много мест, где можно положить корень. Для разветвления помеченных деревьев общее количество корневых деревьев составляет:
 для
для  ,
,  представляет количество листовых узлов.
представляет количество листовых узлов.Для разветвленных помеченных деревьев общее количество деревьев без корней составляет:
 для
для  .
.Среди помеченных ветвящихся деревьев количество неукорененных деревьев с 
Количество корневых деревьев быстро растет в зависимости от количества кончиков. Для 10 подсказок существует более 
| Помеченные. листья | Бинарные. деревья без корней | Бинарные. деревья с корнями | Многоцветковые. деревья с корнями | Все возможные. корневые деревья |
|---|---|---|---|---|
| 1 | 1 | 1 | 0 | 1 |
| 2 | 1 | 1 | 0 | 1 |
| 3 | 1 | 3 | 1 | 4 |
| 4 | 3 | 15 | 11 | 26 |
| 5 | 15 | 105 | 131 | 236 |
| 6 | 105 | 945 | 1,807 | 2,752 |
| 7 | 945 | 10,395 | 28,813 | 39,208 |
| 8 | 10,395 | 135,135 | 524,897 | 660,032 |
| 9 | 135,135 | 2,027,025 | 10,791,887 | 12,818,912 |
| 10 | 2,027,025 | 34,459,425 | 247,678,399 | 282,137,824 |
 Дендрограмма филогении некоторых пород собак
Дендрограмма филогении некоторых пород собак A дендрограмма - общее название дерева, независимо от того, филогенетический или нет, и, следовательно, также для схематического представления филогенетического дерева.
A кладограмма представляет только образец ветвления; т. е. длина его ветвей не отражает время или относительную величину изменения символа, а его внутренние узлы не представляют предков.
Филограмма - это филогенетическое дерево, длина ветвей которого пропорциональна количество изменения символа.
 Хронограмма чешуекрылых. В этом типе филогенетического дерева длины ветвей пропорциональны геологическому времени.
Хронограмма чешуекрылых. В этом типе филогенетического дерева длины ветвей пропорциональны геологическому времени. Хронограмма - это филогенетическое дерево, которое явно представляет время через длину ветвей.
A Дальгренограмма представляет собой диаграмму, представляющую поперечное сечение филогенетического дерева
A филогенетическая сеть строго говоря не дерево, а скорее более общий граф или направленный ациклический граф в случае корневых сетей. Они используются для преодоления некоторых из ограничений, присущих деревьям.
 Диаграмма веретена, показывающая эволюцию позвоночных на уровне класса, ширина веретена указывает количество семейств. Диаграммы веретена часто используются в эволюционной таксономии.
Диаграмма веретена, показывающая эволюцию позвоночных на уровне класса, ширина веретена указывает количество семейств. Диаграммы веретена часто используются в эволюционной таксономии.Диаграмма веретена или пузырьковая диаграмма часто называется ромерограммой после ее популяризации американским палеонтологом Альфредом Ромером. Он представляет таксономическое разнообразие (горизонтальная ширина) по сравнению с геологическим временем (вертикальная ось), чтобы отразить изменение численности различных таксонов во времени. Однако диаграмма веретена не является эволюционным деревом: таксономические веретена скрывают фактические отношения родительского таксона с дочерним таксоном и имеют недостаток, заключающийся в вовлечении парафилии родительской группы. Этот тип диаграммы больше не используется в первоначально предложенной форме.
 Коралл жизни
Коралл жизни Дарвин также упомянул, что коралл может быть более подходящей метафорой, чем дерево. Действительно, филогенетические кораллы полезны для изображения прошлой и настоящей жизни, и у них есть некоторые преимущества перед деревьями (разрешены анастомозы и т. Д.).
Составленные филогенетические деревья с нетривиальным количеством входных последовательностей построены с использованием методов вычислительной филогенетики. Методы матрицы расстояний, такие как соединение соседей или UPGMA, которые вычисляют генетическое расстояние из множественных выравниваний последовательностей, наиболее просты в реализации, но не используйте эволюционную модель. Многие методы выравнивания последовательностей, такие как ClustalW, также создают деревья с использованием более простых алгоритмов (т.е. основанных на расстоянии) построения дерева. Максимальная экономия - еще один простой метод оценки филогенетических деревьев, но подразумевает неявную модель эволюции (то есть экономию). Более продвинутые методы используют критерий оптимальности максимального правдоподобия, часто в рамках байесовской структуры, и применяют явную модель эволюции для оценки филогенетического дерева. Определение оптимального дерева с использованием многих из этих методов является NP-трудным, поэтому методы эвристического поиска и оптимизации используются в сочетании с функциями оценки дерева для определения достаточно хорошее дерево, которое соответствует данным.
Методы построения дерева можно оценить по нескольким критериям:
Методы построения деревьев также привлекли внимание математиков. Деревья также могут быть построены с использованием T-theory.
Деревья могут быть закодированы в различных форматах, каждый из которых должен представлять вложенную структуру дерева. Они могут кодировать, а могут и не кодировать длины ветвей и другие особенности. Стандартизированные форматы имеют решающее значение для распространения и совместного использования деревьев, не полагаясь на вывод графики, который трудно импортировать в существующее программное обеспечение. Обычно используются следующие форматы:
Хотя филогенетические деревья созданы на основе секвенированных генов или геномных данные по разным видам могут дать представление об эволюции, этот анализ имеет важные ограничения. Что наиболее важно, деревья, которые они производят, не обязательно правильные - они не обязательно точно отражают эволюционную историю включенных таксонов. Как и любые научные результаты, они могут быть фальсифицированы в результате дальнейшего изучения (например, сбора дополнительных данных, анализа существующих данных улучшенными методами). Данные, на которых они основаны, могут быть зашумленными ; анализ может быть затруднен из-за генетической рекомбинации, горизонтального переноса генов, гибридизации между видами, которые не были ближайшими соседями на дереве до того, как произойдет гибридизация, конвергентная эволюция и консервативные последовательности.
Кроме того, существуют проблемы, связанные с основанием анализа на одном типе признака, таком как единственный ген или белок или только на основе морфологического анализа, потому что такие деревья, построенные из другого несвязанного источника данных, часто отличаются от первого, и поэтому требуется большая осторожность при выводе филогенетических отношений между видами. Это наиболее верно в отношении генетического материала, который подвержен латеральному переносу генов и рекомбинации, где разные блоки гаплотипа могут иметь разную историю. В этих типах анализа выходное дерево филогенетического анализа отдельного гена является оценкой филогении гена (т. Е. Генного дерева), а не филогении таксонов (т. Е. Древа видов), из которых эти символы были выбраны, хотя в идеале оба должны быть очень близки. По этой причине серьезные филогенетические исследования обычно используют комбинацию генов, происходящих из разных геномных источников (например, из митохондриальных или пластидных или ядерных геномов), или генов, которые, как ожидается, будут развиваться при различных режимах отбора, так что гомоплазия (ложная гомология ) вряд ли возникнет в результате естественного отбора.
Когда вымершие виды включены в анализ в качестве конечных узлов (а не, например, для ограничения внутренних узлов), они считаются не представляющими прямых предков каких-либо существующих видов. Вымершие виды обычно не содержат высококачественной ДНК.
Диапазон полезных материалов ДНК расширился с развитием технологий экстракции и секвенирования. Разработка технологий, способных выводить последовательности из более мелких фрагментов или из пространственных структур продуктов деградации ДНК, еще больше расширит диапазон ДНК, считающихся полезными.
Филогенетические деревья также могут быть выведены из ряда других типов данных, включая морфологию, наличие или отсутствие определенных типов генов, события вставки и удаления - и любое другое наблюдение, которое, как считается, содержит эволюционный сигнал.
Филогенетические сети используются, когда разветвляющиеся деревья не подходят из-за этих сложностей, которые предполагают более сетчатую эволюционную историю отобранных организмов.
| Викискладе есть медиафайлы, связанные с Филогенетическими деревьями. |