
Войти
 Гомологичные hox-гены у таких разных животных, как насекомые и позвоночные, контролируют эмбриональное развитие и, следовательно, форму взрослых тел. Эти гены были высококонсервативными на протяжении сотен миллионов лет эволюции.
Гомологичные hox-гены у таких разных животных, как насекомые и позвоночные, контролируют эмбриональное развитие и, следовательно, форму взрослых тел. Эти гены были высококонсервативными на протяжении сотен миллионов лет эволюции.Эволюционная биология развития (неформально evo-DevO ) - это область биологические исследования, которые сравнивают процессы развития различных организмов с делают вывод о родственных отношениях между ними и о том, как процессы развития эволюционировал.
Эта область росла с начала XIX века, когда эмбриология столкнулась с загадкой: зоологи не знали, как эмбриональное развитие контролировалось в молекулярный уровень. Чарльз Дарвин отметил, что наличие похожих эмбрионов подразумевает общее происхождение, но до 1970-х годов не было достигнуто большого прогресса. Затем технология рекомбинантной ДНК наконец-то объединила эмбриологию с молекулярной генетикой. Ключевым ранним открытием стали гомеотические гены, которые регулируют развитие широкого круга эукариот.
. Эта область характеризуется некоторыми ключевыми концепциями, которые застали биологов-эволюционистов врасплох. Один из них - глубокая гомология, открытие, что разные органы, такие как глаза насекомых, позвоночных и головоногих моллюсков, долгое время считались развивающиеся отдельно, контролируются аналогичными генами, такими как pax-6, из набора гена evo-Dev. Эти гены являются древними и высококонсервативными среди филы ; они генерируют модели во времени и пространстве, которые формируют эмбрион, и в конечном итоге формируют план тела организма. Другая причина заключается в том, что виды не сильно различаются по своим структурным генам, например генам, кодирующим ферменты ; что действительно отличается, так это способ, которым экспрессия гена регулируется с помощью генов инструментария. Эти гены повторно используются, без изменений, много раз в разных частях эмбриона и на разных стадиях развития, образуя сложный каскад контроля, включая и выключая другие регуляторные гены, а также структурные гены по точному шаблону. Это многократное плейотропное повторное использование объясняет, почему эти гены являются высококонсервативными, поскольку любое изменение будет иметь множество неблагоприятных последствий, которым естественный отбор будет противодействовать.
Новые морфологические признаки и, в конечном итоге, новые виды создаются вариациями в наборе инструментов, либо когда гены экспрессируются в новом образце, либо когда гены набора инструментов приобретают дополнительные функции. Другой возможностью является неоламаркистская теория, согласно которой эпигенетические изменения позже консолидируются на генном уровне, что, возможно, имело важное значение в начале истории многоклеточной жизни.
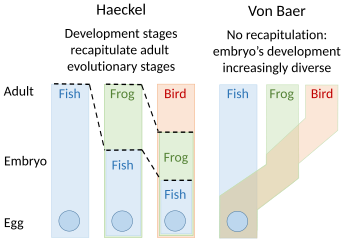 Теории эмбриологии из Эрнста Геккеля, который выступал за рекапитуляцию эволюционного развития эмбриона, и эпигенез Карла Эрнста фон Бэра
Теории эмбриологии из Эрнста Геккеля, который выступал за рекапитуляцию эволюционного развития эмбриона, и эпигенез Карла Эрнста фон Бэра A Теория перепросмотра эволюционного развития была предложена Этьеном Серром в 1824–1826 годах, вторя идеям 1808 года Иоганна Фридриха Меккеля. Они утверждали, что эмбрионы «высших» животных прошли или повторяли серию стадий, каждая из которых напоминала животное, находящееся ниже по великой цепи бытия. Например, мозг человеческого эмбриона сначала выглядел как мозг рыбы, а затем, в свою очередь, как мозг рептилии, птицы и млекопитающее, прежде чем стать явно человеком. Эмбриолог Карл Эрнст фон Бэр возражал против этого, утверждая в 1828 году, что не существует линейной последовательности, как в великой цепи бытия, основанной на едином плане тела, а есть процесс эпигенез, в котором дифференцируются структуры. Фон Баер вместо этого распознал четыре различных плана тела животных : излучение, как морская звезда ; моллюск, например моллюск ; членораздельно, как лобстеры ; и позвоночные, такие как рыба. Тогда зоологи в значительной степени отказались от перепросмотра, хотя Эрнст Геккель возродил его в 1866 году.
 А. Ланцетник (хордовая), B. Личиночная оболочка, C. Взрослая оболочка. Ковалевский увидел, что хорда (1) и жаберная щель (5) являются общими для оболочников и позвоночных.
А. Ланцетник (хордовая), B. Личиночная оболочка, C. Взрослая оболочка. Ковалевский увидел, что хорда (1) и жаберная щель (5) являются общими для оболочников и позвоночных. С начала 19 века до большей части 20 века Эмбриология столкнулась с загадкой. Было замечено, что животные из яйца развиваются во взрослых строения тела, часто через похожие стадии, но зоологи почти ничего не знали о том, как эмбриональное развитие контролировалось в на молекулярном уровне, и поэтому столь же мало о том, как процессы развития эволюционировали. Чарльз Дарвин утверждал, что общая эмбриональная структура подразумевает общего предка. В качестве примера Дарвин в своей книге 1859 года О происхождении видов привел креветочную -подобную личинку ракушка, чьи сидячие взрослые особи не были похожи на других членистоногих ; Линней и Кювье классифицировали их как моллюсков. Дарвин также отметил открытие Александра Ковалевского, что оболочка тоже не была моллюском, но на личиночной стадии имела хорду и образовавшиеся глоточные щели. из тех же зародышевых листков, что и эквивалентные структуры у позвоночных, и поэтому их следует сгруппировать с ними как хордовые. Таким образом, зоология XIX века превратила эмбриологию в эволюционную науку, соединив филогению с гомологиями зародышевых листков зародыша. Зоологи, в том числе Фриц Мюллер, предложили использовать эмбриологию для обнаружения филогенетических отношений между таксонами. Мюллер продемонстрировал, что ракообразные имеют общую личинку науплиуса, определив несколько паразитических видов, которые не были признаны ракообразными. Мюллер также признал, что естественный отбор должен воздействовать на личинок точно так же, как и на взрослых особей, опровергая перепросмотр, который потребовал бы защиты личиночных форм от естественного отбора. Две другие идеи Геккеля об эволюции развития оправдались лучше, чем перепросмотр: в 1870-х годах он утверждал, что изменения во времени (гетерохрония ) и изменения положения в теле (гетеротопия ) аспектов эмбрионального развития будет стимулировать эволюцию, изменив форму тела потомка по сравнению с телом предка. Прошло столетие, прежде чем эти идеи оказались верными. В 1917 году Д'Арси Томпсон написал книгу о формах животных, показав с помощью простой математики, как небольшие изменения параметров, такие как углы спиральной раковины брюхоногого моллюска, могут радикально изменить форму животного, хотя он предпочитал механическое объяснение эволюционному объяснению. Но в следующем столетии без молекулярных доказательств прогресс застопорился.
В так называемом современном синтезе начала 20-го века столетия, Рональд Фишер объединил теорию Дарвина эволюции с его настойчивостью на естественном отборе, вариации и Грегора Менделя. законы генетики в целостную структуру для эволюционной биологии. Биологи предположили, что организм является прямым отражением составляющих его генов: генов, кодирующих белки, из которых строится тело организма. Биохимические пути (и, как они предполагали, новые виды) эволюционировали в результате мутаций в этих генах. Это была простая, ясная и почти полная картина: но она не объясняла эмбриологию.
Эволюционный эмбриолог Гэвин де Бир предвосхитил эволюционную биологию развития в своей книге 1930 года Эмбрионы и предки, показывая, что эволюция могла происходить за счет гетерохронии, например, в сохранения ювенильных черт у взрослого. Это, как утверждал де Бир, могло вызвать очевидные внезапные изменения в летописи окаменелостей , поскольку эмбрионы плохо окаменелости. Поскольку пробелы в летописи окаменелостей использовались в качестве аргумента против постепенной эволюции Дарвина, объяснение де Бера поддерживало дарвиновскую позицию. Однако, несмотря на де Бера, современный синтез в значительной степени игнорировал эмбриональное развитие для объяснения формы организмов, поскольку популяционная генетика оказалась адекватным объяснением того, как развивались формы.
 лак оперон. Вверху: подавленный, внизу: активный. 1: РНК-полимераза, 2: Репрессор, 3: Промотор, 4: оператор, 5: Лактоза, 6–8: гены, кодирующие белок, контролируемые переключателем, которые вызывают переваривание лактозы
лак оперон. Вверху: подавленный, внизу: активный. 1: РНК-полимераза, 2: Репрессор, 3: Промотор, 4: оператор, 5: Лактоза, 6–8: гены, кодирующие белок, контролируемые переключателем, которые вызывают переваривание лактозы В 1961 году Жак Моно, Жан -Пьер Чанжукс и Франсуа Жакоб обнаружили лак-оперон в бактерии Escherichia coli. Это был кластер генов, скомпонованных в контур управления с обратной связью , так что его продукты производились только при "включении" стимулом окружающей среды. Одним из этих продуктов был фермент, расщепляющий сахар, лактозу; и сама лактоза была стимулом, который включал гены. Это было откровением, поскольку впервые было показано, что гены даже в таком маленьком организме, как бактерия, подлежат точному контролю. Подразумевалось, что многие другие гены также тщательно регулируются.
В 1977 году началась революция в представлениях об эволюции и биологии развития, с появлением появление технологии рекомбинантной ДНК в генетике, а также работы Стивена Дж. Гулда «Онтогенез и филогения» и Франсуа Джейкоба. Гулд положил конец интерпретации Геккеля эволюционной эмбриологии, а Якоб изложил альтернативную теорию. Это привело к второму синтезу, включающему наконец эмбриологию, а также молекулярную генетику, филогению и эволюционную биологию, чтобы сформировать evo-DevO. В 1978 году Эдвард Б. Льюис открыл гомеотические гены, регулирующие эмбриональное развитие у плодовых мух Drosophila, которые, как и все насекомые, являются членистоногими, один из основных типов беспозвоночных животных. Билл МакГиннис быстро обнаружил последовательности гомеотических генов, гомеобоксы, у животных других типов, у позвоночных такие как лягушки, птицы и млекопитающие ; позже они были также обнаружены в грибах, таких как дрожжи, и в растениях. Очевидно, было сильное сходство в генах, контролирующих развитие у всех эукариот. В 1980 году Кристиан Нюсслейн-Фольхард и Эрик Вишаус описали гены пробела, которые помогают создать паттерн сегментации в эмбрионах плодовой мухи ; они и Льюис получили Нобелевскую премию за свою работу в 1995 году.
Позже были обнаружены более специфические сходства: например, ген без дистальных отделов был обнаружен в 1989: участие в развитии придатков или конечностей у плодовых мух, плавников рыб, крыльев кур, параподий морских кольчатых червей, ампул и сифонов оболочников., и трубчатые футы морских ежей. Было очевидно, что ген должен быть древним, восходящим к последнему общему предку двусторонних животных (до эдиакарского периода, который начался около 635 миллионов лет назад). Эво-дево начал раскрывать способы, которыми все тела животных были построены в процессе развития.
Яйца разных животных сферической формы дают поднимаются до самых разных тел, от медуз до омаров, от бабочек до слонов. Многие из этих организмов имеют одни и те же структурные гены белков для построения тела, таких как коллаген и ферменты, но биологи ожидали, что каждая группа животных будет иметь свои собственные правила развития. Удивление evo-DevO заключается в том, что формирование тел контролируется довольно небольшим процентом генов, и что эти регуляторные гены являются древними, общими для всех животных. У жирафа нет гена длинной шеи, как и у слона нет гена большого тела. Их тела формируются системой переключения, которая заставляет развитие различных функций начинаться раньше или позже, происходить в той или иной части эмбриона и продолжаться более или менее долго.
Загадка вопрос о том, как контролировалось эмбриональное развитие, начал решаться с использованием плодовой мушки Drosophila melanogaster в качестве модельного организма. Пошаговый контроль его эмбриогенеза визуализировали путем присоединения флуоресцентных красителей разного цвета к конкретным типам белка, производимого генами, экспрессируемыми в эмбрионе. Краситель, такой как зеленый флуоресцентный белок, первоначально полученный от медузы, обычно был присоединен к антителу, специфичному к белку плодовой мухи, образуя точный индикатор того, где и когда этот белок появился в живом эмбрионе.
 Ген pax-6 контролирует развитие глаз разных типов в животном мире.
Ген pax-6 контролирует развитие глаз разных типов в животном мире. Используя такую технику, в 1994 Уолтер Геринг обнаружил, что ген pax-6, жизненно важный для формирования глаз у плодовых мушек, точно соответствует гену формирования глаз у мышей и людей. Тот же ген был быстро обнаружен у многих других групп животных, таких как кальмар, головоногие моллюски моллюски. Биологи, в том числе Эрнст Майр, считали, что глаза возникали в животном мире по крайней мере 40 раз, так как анатомия разных типов глаз сильно различается. Например, сложный глаз плодовой мушки состоит из сотен небольших линзированных структур (ommatidia ); человеческий глаз имеет слепое пятно, где зрительный нерв входит в глаз, а нервные волокна проходят по поверхности сетчатки, поэтому свет должен пройти через слой нервных волокон, прежде чем достигнет детекторных клеток в сетчатке, так что структура фактически является «перевернутой»; напротив, у головоногого глаза есть сетчатка, затем слой нервных волокон, а затем стенка глаза «на правильном пути». Однако доказательства pax-6 заключались в том, что одни и те же гены контролировали развитие глаз всех этих животных, что позволяет предположить, что все они произошли от общего предка. Древние гены были сохранены посредством миллионы лет эволюции для создания разнородных структур для аналогичных функций, демонстрирующих глубокую гомологию между структурами, которые когда-то считались чисто аналогичными. Это понятие позже было распространено на эволюцию эмбриогенеза и привело к радикальному пересмотру значения гомологии в эволюционной биологии.
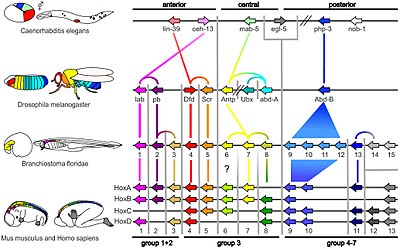
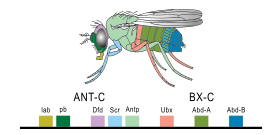 Экспрессия гомеобокса (Hox) гены плодовой мухи
Экспрессия гомеобокса (Hox) гены плодовой мухи Небольшая часть генов в геноме организма контролирует развитие организма. Эти гены называются генетически-развитым инструментарием. Они высококонсервативны среди типов и максимально экспрессируются на филотипической стадии, что означает, что они древние и очень похожи у широко разделенных групп животных. Различия в развертывании генов набора инструментов влияют на план тела и количество, идентичность и структуру частей тела. Большинство генов инструментария являются частями сигнальных путей : они кодируют факторы транскрипции, белки клеточной адгезии, белки рецептора на поверхности клетки и сигнальные лиганды, которые связываются с ними, и секретируют морфогены, которые диффундируют через эмбрион. Все это помогает определить судьбу недифференцированных клеток эмбриона. Вместе они создают модели во времени и пространстве, которые формируют эмбрион, и в конечном итоге формируют план тела организма. Среди наиболее важных генов инструментария - Hox-гены. Эти факторы транскрипции содержат мотив ДНК, связывающий белок гомеобокс, также обнаруживаемый в других генах инструментального набора, и создают основной рисунок тела вдоль его оси спереди назад. Hox-гены определяют, где повторяющиеся части, такие как множество позвонков змей, будут расти в развивающемся эмбрионе или личинке. Pax-6, как уже упоминалось, является классическим геном инструментария. Хотя другие гены набора участвуют в создании bodyplan растения, гены гомеобокса также встречаются у растений, что означает, что они являются общими для всех эукариот.
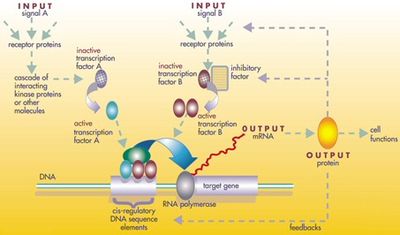 A регуляторная сеть генов
A регуляторная сеть генов Белковые продукты регуляторного инструментария повторно используются не путем дупликации и модификации, а путем сложной мозаики плейотропии, применяемой без изменений во многих независимых процессах развития, давая образец для многих разное строение тела. Локусы этих генов плейотропного инструментария имеют большие, сложные и модульные цис-регуляторные элементы. Например, в то время как неплейотропный ген родопсина у плодовой мушки имеет цис-регуляторный элемент длиной всего несколько сотен пар оснований, плейотропный безглазый цис -регуляторная область содержит 6 цис-регуляторных элементов в более чем 7000 пар оснований. Вовлеченные регулирующие сети часто бывают очень большими. Каждый регуляторный белок контролирует «до сотен» цис-регуляторных элементов. Например, 67 факторов транскрипции плодовой мушки контролируют в среднем 124 гена-мишени каждый. Вся эта сложность позволяет генам, участвующим в развитии эмбриона, включаться и выключаться точно в нужное время и точно в нужных местах. Некоторые из этих генов являются структурными, непосредственно образующими ферменты, ткани и органы эмбриона. Но многие другие сами по себе являются регуляторными генами, поэтому часто включается точно рассчитанный по времени каскад переключения, включающий включение одного процесса развития за другим в развивающемся эмбрионе.
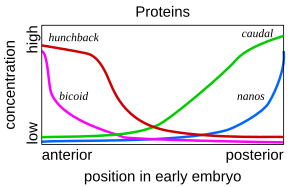 Распределение генных продуктов по длинной оси раннего эмбриона плодовой мухи
Распределение генных продуктов по длинной оси раннего эмбриона плодовой мухи Такая каскадная регуляторная сеть была подробно изучена при развитии эмбриона плодовой мухи. Молодой эмбрион имеет овальную форму, похожую на мяч для регби. Небольшое количество генов продуцирует информационные РНК, которые создают градиенты концентрации вдоль длинной оси эмбриона. В раннем эмбрионе гены bicoid и hunchback находятся в высокой концентрации около переднего конца и определяют структуру будущей головы и грудной клетки; каудальный гены и гены nanos находятся в высокой концентрации около заднего конца и определяют структуру самых задних сегментов брюшка. Эффекты этих генов взаимодействуют; например, белок Bicoid блокирует трансляцию информационной РНК каудального отдела, поэтому концентрация каудального белка становится низкой на переднем конце. Позже каудальный включает гены, которые создают самые задние сегменты мухи, но только на заднем конце, где он наиболее сконцентрирован.
 Гены пробела у плодовой мухи включаются такими генами, как bicoid, формируя полосы на эмбрионе, которые начинают формировать узор сегментов тела.
Гены пробела у плодовой мухи включаются такими генами, как bicoid, формируя полосы на эмбрионе, которые начинают формировать узор сегментов тела. Белки Bicoid, Hunchback и Caudal, в свою очередь, регулируют транскрипцию генов гэпа, таких как гигантский, книрпс, Krüppel и бесхвостый полосатый узор, создающий первый уровень структур, которые станут сегментами. Белки из них, в свою очередь, контролируют гены парных правил , которые на следующем этапе создают 7 полос поперек длинной оси эмбриона. Наконец, гены полярности сегментов, такие как engrailed, разделяют каждую из 7 полос на две, создавая 14 будущих сегментов.
Этот процесс объясняет точную консервацию последовательностей генов набора инструментов, что привело к глубокая гомология и функциональная эквивалентность белков инструментария у разнородных животных (видно, например, когда белок мыши контролирует развитие плодовой мухи). Взаимодействие факторов транскрипции и цис-регуляторных элементов или сигнальных белков и рецепторов блокируется из-за многократного использования, что делает почти любую мутацию вредной и, следовательно, устраняется естественным отбором.
Среди наиболее удивительных и, возможно, противоречивых (с неодарвинистской точки зрения) результатов недавних исследований в области эволюционной биологии развития является то, что разнообразие строений тела и морфология организмов многих типов не обязательно отражается в разнообразии на уровне последовательностей генов, включая последовательности генетического инструментария развития и других генов, участвующих в развитии. Действительно, как отметили Джон Герхарт и Марк Киршнер, существует очевидный парадокс: «там, где мы больше всего ожидаем найти вариацию, мы находим сохранение, отсутствие изменений». Итак, если наблюдаемая морфологическая новизна между различными кладами не является результатом изменений в последовательностях генов (например, в результате мутации ), откуда она взялась? Новизна может возникнуть в результате мутационных изменений в регуляции гена.
 Heliconius erato
Heliconius erato  Heliconius melpomene Различные виды бабочки Heliconius независимо развили сходные модели, очевидно, как , так и ограниченные доступным онтогенетическим набором инструментов гены, контролирующие формирование паттерна крыла .
Heliconius melpomene Различные виды бабочки Heliconius независимо развили сходные модели, очевидно, как , так и ограниченные доступным онтогенетическим набором инструментов гены, контролирующие формирование паттерна крыла .Вариации в наборе инструментов, возможно, повлияли на большую часть морфологической эволюции животных. Этот инструментарий может стимулировать эволюцию двумя способами. Ген набора инструментов может быть выражен по-другому, например, когда клюв Дарвина большого земляного вьюрка был увеличен геном BMP, или когда змеи потеряли свои ноги, как бесконечно удаленные. стали недостаточно выраженными или совсем не выраженными там, где другие рептилии продолжали формировать свои конечности. Или ген набора инструментов может приобрести новую функцию, как видно из множества функций того же гена, без дистального отдела, который контролирует такие разнообразные структуры, как нижняя челюсть у позвоночных, ноги и усики у плодовой мухи и глазное пятно узор в бабочка крылья. Учитывая, что небольшие изменения в генах набора инструментов могут вызывать значительные изменения в структурах тела, они часто включают одну и ту же функцию конвергентно или параллельно. distal-less формирует рисунок крыльев у бабочек Heliconius erato и Heliconius melpomene, которые имитируют Мюллера. В так называемой облегченной вариации их крылья возникли в ходе разных эволюционных событий, но контролируются одними и теми же генами. Изменения развития могут напрямую способствовать видообразованию.
Эволюционные инновации иногда могут начинаться в ламарковском стиле с эпигенетических изменений регуляции генов или поколение фенотипа, впоследствии закрепленное изменениями на уровне гена. Эпигенетические изменения включают модификацию ДНК путем обратимого метилирования, а также незапрограммированное изменение формы организма под действием физических и других воздействий окружающей среды из-за присущей пластичности механизмов развития. Биологи Стюарт А. Ньюман и Герд Б. Мюллер предположили, что организмы на раннем этапе истории многоклеточной жизни были более восприимчивы к этой второй категории эпигенетической детерминации, чем современные организмы. основа для ранних макроэволюционных изменений.
 Среди многоножек все представители геофиломорф ограничены развитием предвзятость иметь нечетное количество сегментов, всего от 27 до 191.
Среди многоножек все представители геофиломорф ограничены развитием предвзятость иметь нечетное количество сегментов, всего от 27 до 191. Развитие в конкретных линиях может быть смещено либо положительно, в сторону данной траектории или фенотипа, либо отрицательно, не вызывая определенных типов изменений ; либо может быть абсолютным (изменение всегда или никогда не происходит), либо относительным. Однако доказательства любого такого направления эволюции получить сложно, и они также могут быть результатом ограничений развития, которые ограничивают диверсификацию. Например, у брюхоногих моллюсков раковина улитки всегда построена как трубка, которая увеличивается как в длину, так и в диаметре; Выбор создал широкий спектр форм оболочки, таких как плоские спирали, каури и высокие спирали револьверной головки в рамках этих ограничений. Среди многоножек, Lithobiomorpha всегда имеют 15 сегментов туловища во взрослом возрасте, вероятно, в результате смещения развития в сторону нечетного количества сегментов туловища. Другой отряд сороконожек, Geophilomorpha, количество сегментов варьируется у разных видов от 27 до 191, но число всегда нечетное, что делает это абсолютным ограничением; почти все нечетные числа в этом диапазоне заняты тем или иным видом.
Экологическая эволюционная биология развития (eco-evo-Dev ) объединяет исследования биологии развития и экологии для изучения их взаимосвязи с теорией эволюции. Исследователи изучают такие концепции и механизмы, как пластичность развития, эпигенетическая наследственность, генетическая ассимиляция, построение ниши и симбиоз.