
Войти
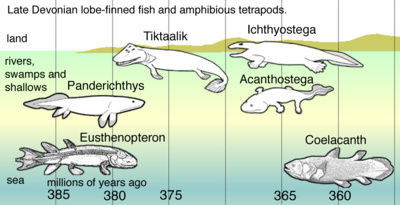 В позднем девоне позвоночных видообразование, потомки пелагических рыб с лопастными плавниками - например, Eusthenopteron - применяли последовательность адаптаций: * Panderichthys, подходящих для илистые отмели * Тиктаалик с плавниками, похожими на конечности, которые могли перенести его насушу * Ранние четвероногие в заросших сорняках болотах, например: ** Acanthostega, у которых были ноги с восемью пальцами * * Ichthyostega с конечностями. Потомки также включаются пелагических рыб с лопастными плавниками, таких как виды целакантов.
В позднем девоне позвоночных видообразование, потомки пелагических рыб с лопастными плавниками - например, Eusthenopteron - применяли последовательность адаптаций: * Panderichthys, подходящих для илистые отмели * Тиктаалик с плавниками, похожими на конечности, которые могли перенести его насушу * Ранние четвероногие в заросших сорняках болотах, например: ** Acanthostega, у которых были ноги с восемью пальцами * * Ichthyostega с конечностями. Потомки также включаются пелагических рыб с лопастными плавниками, таких как виды целакантов. Эволюция четвероногих началась около 400 миллионов лет назад в Девонский период с самыми ранними четвероногими произошел от рыб с лопастными плавниками. Тетраподы (в соответствиис определением на основе апоморфии, используемым на этой странице) к категории животных в биологическом суперклассе Tetrapoda, который включает всех живых и вымерших амфибии, рептилии, птицы и млекопитающие. В то время как большинство современных видов наземных, мало доказательств поддерживает идею о том, что какие-либо из самых ранних источников передвигаться по суше, поскольку их конечности не могли установить свое телоот земли, а следы не указаны на то, что они волочили живот. Предположительно, следы животных, идущие по дну неглубоких водоемов. Конкретные водные предки четвероногих и процесс колонизации суши неясными и в настоящее время являются областью активных исследований и дискуссий среди палеонтологов.
Большинство земноводных сегодня остаются полуводными, проживая первую стадию своей жизни как рыбоподобные головастики. Несколько групп четвероногих, таких как змеи и китообразные, потеряли некоторые или все свои конечности. Кроме того, многие четвероногие вернулись к частично водной или полностью водной жизни на протяжении всей истории группы (современные примеры полностью водных четвероногих включают китообразных и сирен ). Первое возвращение к водному образу жизни могло произойти уже в каменноугольном периоде, тогда как другое возвращение произошло совсем недавно, в кайнозое, как у китообразных ластоногих, и несколько современных амфибий.
Переход от плана для дыхания и навигации в воде к плану тела, позволяющему животному перемещаться по суше, является одним из самых глубоких известных изменений. Он также является одним из наиболее изученных, во многом ряду значительных переходных ископаемых находок в конце 20 века в сочетании с улучшенным филогенетическим анализом.
Девонский период традиционно известен как «Эпоха рыб», указая диверси задачи вымерших и современных крупных рыб. группы. Среди них были ранние костистые рыбы, которые в начале периода разнообразили и распространились в пресноводных и солоноватых средах. Ранние типы напоминали своих хрящевых предков вомногих чертах анатомии, включая акулоподобный хвостовой плавник, спиральную кишку, большие грудные плавники, укрепленные спереди скелетными элементами, и в основном неокостеневший осевой скелет.
них были черты, отделяющие их от хрящевых рыб, черты, которые стали ключевыми в эволюции земных форм. За исключением пары дыхательные пути, жабры не открываются наружу по отдельной, как у акул; скорее, они были заключены в жаберную камеру, укрепленную мембранными костями и покрытую костной operculum с единственным отверстием наружу. Клейтрум, образующий задний край жаберной камеры, также служил якорем для грудных плавников. У хрящевых рыб такого крепления для грудных плавников нет. Это может быть конструкция конструкции у четвероногих, позже она будет использована в конструкции конструкции у ранних костистых рыб. Как часть общей брони ромбовидной космической чешуи, череп полностью покрыткожной костью, составляющая крыша черепа над акулоподобным хрящом внутренним черепом. Важно отметить, что у них также была пара вентральных парных легких, чего не хватало у акул и скатов.
Предполагалось, что рыбы в степени эволюционировали вокруг рифов, но с момента своего возникновения 480 миллионов лет назад они жили в прибрежных средах, таких как приливные зоны или постоянно мелководные лагуны и не начали распространяться в другиебиотопы раньше 60 миллионов лет спустя. Некоторые приспособились к более глубокой воде, в то время как твердые и массивные формы остались там, где, или мигрировали в пресную воду. Увеличение первичной продуктивности на суше в конце девона изменило пресноводные экосистемы. Когда питательные вещества из растений попадали в озеро и реки, они поглощались микроорганизмами, которые, в свою очередь, поедались беспозвоночными, которые служили пищей для позвоночных. Некоторые рыбы также стали детритофагами. Ранние четвероногие развили толерантность к средам с различной соленостью, такой какья или дельты.
Легкие / плавательный пузырь возник как вырост из кишечника, образуя газовый наполненный мочевой пузырь над пищеварительной системой. В своей примитивной форме воздушный пузырь был открыт в пищеварительный канал, состояние, называемое физостом, которое до сих пор встречается у многих у многих. Основная функция не совсемопределена. Одно из соображений - плавучесть. Тяжелые чешуйчатые доспехи ранних костистых рыб, несомненно, утяжеляют животных. У хрящевых рыб, лишенных плавательного пузыря, акулам открытого моря необходимо постоянно плавать, чтобы не провалиться на глубину, а грудные плавники обеспечивают подъемную силу. Другой фактор - потребление кислорода. Кислород в окружающей среде в раннем девоне был относительно низко, возможно, около половины современных значений. Животные свысокими потребностями энергии по сравнению с беспозвоночными аналогичного размера. В девоне наблюдалось повышение уровня кислорода, что открыло новые экологические ниши, группам людей использовать дополнительный кислород для активных, крупных животных. В частности, в местах обитания на тропических болотах атмосферный кислород более стабилен и, возможно, заставил полагаться на легкие, а не на жабры первичного кислорода. В конце концов, плавучесть и дыхание, возможно, были важны, и некоторыесовременные физиостомные рыбы действительно используют свой мочевой пузырь для обоих.
Для газообмена легким требуется кровоснабжение. У хрящевых рыб и костистых рыб сердце расположено низко в теле и перекачивает кровь вперед через брюшную аорту, которая разделяется на ряд парных дуг аорты., каждую из которых соответствует жаберной дуге. Затем дуги аорты сливаются над жабрами, образуя дорсальную аорту, снабжает насыщенной кислородомкровью. У двоякодышащих, боуфинов и бичиров плавательный пузырь снабжается кровью парными легочными артериями, отходящими от самой задней (6-й) дуга аорты. Такой же образец обнаружен у двоякодышащих рыб Protopterus и у наземных саламандр, и, вероятно, он был обнаружен у непосредственных предков четвероногих, а также у первых четвероногих. У многих других костных рыб плавательный пузырь снабжается кровью через спинную аорту.
Для того, чтобы легкие могли осуществлять газообмен, легкие сначала должны иметь в них газ.. У четвероногих от ранних предков современных сохранились три важных механизма дыхания, первым из которых является система обнаружения CO 2 / H +. При дыхании современной тетрапод импульс к вдоху вызывается накоплением CO 2 в кровотоке, а не недостатком O 2. Аналогичная система обнаружения CO 2 / H + обнаружена во всех Osteichthyes, чтоозначает, что последний общий предок всех Osteichthyes нуждался в подобном обнаружении. система. Второй механизм дыхания - это система сурфактанта в легкой, облегчающая газообмен. Это также встречается во всех Osteichthyes, даже в тех, которые почти полностью водные. Высококонсервативный характер системы предполагает, что даже водные Остейхти нуждаются в поверхностно-активных веществах, что может показаться странным, поскольку под водой нет газа. Третий механизм дыхания - этособственное движение дыхания. Этот механизм предшествует последнему общему предку, Остейхтиса, как это можно наблюдать в Лампетра камтшатика, родственной кладе Остейхтиса. У миног этот механизм принимает форму «кашля», когда минога трясет своим телом, позволяя воде течь по жабрам. Когда уровни CO 2 в крови миноги высокие слишком высокие, центральный генератор сигналов посылает сигнал, который заставляет миногу «кашлять» и позволяет CO 2 покинуть свое тело.Эта связь между системой обнаружения CO 2 и центральным генератором паттернов очень похожа на связь между этими двумя системами у четвероногих, что подразумевает гомологию.
ноздри у многих костистых рыб отличаются от ноздрей четвероногих. Обычно у костистых рыб четыре ноздри (носовые отверстия), по одному наре за другим с каждой стороны. Когда рыба плавает, вода течет в переднюю пару, через обонятельную ткань и выходитчерез задние отверстия. Это верно не только для лучеплавниковой рыбы, но и для латимерии, рыбы, включенной в Sarcopterygii, группу, которая также включает четвероногих. Напротив, у четвероногих есть только одна пара ноздрей снаружи, но также есть пара внутренних ноздрей, называемых хоанами, позволяющих им втягивать воздух через нос. Двоякодышащие также являются саркоптеригами с внутренними ноздрями, но они настолько отличаются от хоан четвероногих, что долгое времясчитались независимым развитием.
Эволюция внутренних ноздрей четвероногих горячо обсуждалась в 20 веке. Внутренние ноздри могут быть одним набором внешних (обычно считается задней парой), которые мигрировали в рот, или внутренняя пара может быть недавно сформированной структурой. Однако освободить место для перемещения, две несущие кости верхней челюсти, верхняя челюсть и предчелюстная кость, должны были бы разделиться, чтобы пропустить ноздрю, а снова соединиться. ;до недавнего времени не было свидетельств переходной стадии, когда две кости были разъединены. Такие доказательства теперь доступны: небольшая рыба с лопастными плавниками, названная Kenichthys, найденная в Китае и датированная примерно 395 миллионов лет, представляет собой эволюцию, «пойманную в середине акта», с разделением верхней и челюстей. отверстие - зарождающаяся хоана - на губе между двумя костями. Кенихтис более близок к четвероногим, чем латимерия, у которой есть тольковнешние ноздри; таким образом, он представляет собой промежуточную стадию в развитии состояния четвероногих. Однако причина эволюционного движения задней ноздри от носа к губе до конца не изучена.
 >девонские рыбы, в том числе ранняя акула Cladoselache, Eusthenopteron и рыбы с лопастными плавниками, а также плакодерма ботриолепис (Джозеф Смит, 1905).
>девонские рыбы, в том числе ранняя акула Cladoselache, Eusthenopteron и рыбы с лопастными плавниками, а также плакодерма ботриолепис (Джозеф Смит, 1905). Родственники Кенихтиса вскоре появились водные путях исолоноватых устьях и стали самой многочисленной из костистых рыб на всем протяжении всего Девона и других карбона. Основная анатомия группы хорошо известна благодаря очень подробной работе над Евстеноптерон, выполненной Эриком Ярвиком во второй половине 20 века. Кости крыши черепа были в целом похожи на кости ранних четвероногих, а зубы имели вздутие эмали, подобное таковому у лабиринтодонтов. Парные плавники имели строение с костями, явногомологичными плечевой кости, локтевой кости и радиусу передних плавников и бедренная кость, большеберцовая кость и малоберцовая кость в тазовых плавниках.
Было несколько семейств: Rhizodontida, Canowindridae, Elpistostegidae, Megalichthyidae, Osteolepidae и Tristichopteridae. Большинство из них были рыбами в открытой воде, а некоторые выросли до оченьбольших размеров; взрослые экземпляры достигают нескольких метров в длину. Rhizodontid Rhizodus, по оценкам, вырос до 7 метров (23 фута), что делает его самой крупной из пресноводных рыб.
Хотя большинство из них были рыбами открытой воды, одна группа, Эльпистостегалы , приспособленные к жизни на мелководье. Они развили плоские тела для движения на очень мелководье, а грудные и тазовые плавники взяли на роль основных органов движения. Большинство срединных плавниковисчезло, остался только протоцеркальный хвостовой плавник. Способность дышать атмосферным воздухом. дыхальце стало большим и выдающимся, что рыбам втягивать воздух.
Четвероногие корни уходят в ранний девон тетраподоморф. Примитивные четвероногие животные произошли от остеолепидных тетраподоморфных рыб с лопастными плавниками (саркоптеригиан-кроссоптеригиан) с двухлопастным мозгом в уплощенном черепе. Группа латимерии представляет собой морские саркоптериговы, которые никогда не приобретали эти мелководные приспособления. Саркоптериги, очевидно, взяли разные линии происхождения и соответственно, разделены на две основные группы: Actinistia (включая целаканты) и Rhipidistia (которые включают вымершие линии лопастоперых рыб. И тетраподоморфов).
 Ребристые плавники, подобные таковым у бичиров, люди для передвижения по земле
Ребристые плавники, подобные таковым у бичиров, люди для передвижения по землеСамый старый известный тетраподоморф - Кенихтис из Китая, возрастом около 395 миллионов лет. Двумя самыми ранними тетраподоморфами, датируемыми 380 млн лет, были Гогонас и Пандерихтис. У них были хоаны, и они использовали свои плавники, чтобы перемещаться по приливным каналам и мелководью, забитой мертвыми ветками и гниющими растениями. Их плавники могли быть использованы для прикрепления к растениям или подобным вещам, пока они лежали в засаде длядобычи. Универсальные характеристики четвероногих передних конечностей, которые сгибаются от локтя, и задних конечностей, которые сгибаются назад от колена, вероятно, можно отнести вперед к ранним четвероногим, живущим на мелководье. вода. Окаменелости тазовых костей из Тиктаалика показывают, если они являются репрезентативными для ранних четвероногих в целом, что задние придатки и движение с помощью таза возникли в воде до земной адаптации.
Ещеодно свидетельство того, что ноги и другие черты четвероногих эволюционировали, когда животные были еще водными, так они и питались. У них не было модификаций черепа и челюсти, которые позволяли бы глотать добычу на суше. Добычу можно было поймать на мелководье, у кромки воды или на суше, но ее нужно было поймать в воде, где гидродинамические силы от расширения их ротовой полости заставляли пищу попадать в пищевод.
Это было предположение, что эволюция конечностей четвероногих изплавников рыб с лопастными плавниками связана с экспрессией гена HOXD13 или потерей белков и, которые участвуют в развитии плавников рыб. Моделирование предполагает, что необходимые нервные схемы ходьбы произошли от нервов, управляющих плаванием, с боковыми колебаниями тела, при этом в основном функционируют точки крепления и ограниченная тягу. Этот тип движения, а также изменения грудного пояса, наблюдаются в летописи окаменелостей, могут быть вызваны у бихиров,поднялисьв их воды.
Исследование 2012 года с использованием Трехмерные реконструкции Ихтиостеги пришли к выводу, что он неспособен к типичной четвероногой походке. Конечности не могли двигаться попеременно, так как им не хватало необходимого диапазона вращательных движений. Кроме того, у задних конечностей отсутствовала мускулатура таза, необходимая для движения по земле с помощью задних конечностей. Их наиболее вероятный метод наземного передвижения - этосинхронные «движения с костылями», современные современные прыгунам. Прыгуна показывает, что они движутся, подтягивая себя вперед двумя грудными плавниками одновременно (левый и правый грудные плавники движутся одновременно, а не поочередно). Нет задних "конечностей" / плавников, и значительного изгиба позвоночника.)
Первые четвероногие, вероятно, развились в прибрежных и солоноватых морских средах, а также вмелководных и болотных пресноводных местеобитания. Раньше исследователи считали, что это конец девона. В 2010 году это мнение было оспорено открытием самых старых известных четвероногих, сохранившихся в морских отложениях южного побережья Лавразии, ныне Свентокшиских гор Польши. Они были сделаны на эйфельском этапе в конце среднего девона. Следы, некоторые из которых содержат цифры, датируются примерно 395 миллионами лет назад - на 18миллионов лет раньше, чем самые известные старые окаменелости тела четвероногих. Кроме того, следы показывают, что животное было способно толкать руки и ноги вперед, тип движения, который был бы невозможен для рыб-тетраподоморфов, таких как Тиктаалик. По оценкам, животное, оставившее следы, было до 2,5 (8,2 фута) в длину с подушечками ног до 26 сантиметров (10 дюймов) в ширину, хотя большинство следов имеет длину всего 15 сантиметров (5,9 дюйма).
Новые находки предполагают,что первые четвероногие, возможно, жили как приспособленцы на приливных равнинах, питаясь морскими животными, которые были выброшены на берег или выброшены на берег приливом. Однако в настоящее время рыба в значительном количестве выбрасывается на мель только в определенное время года, как, например, в сезон нереста alewife ; такие высадки не могли обеспечить хищников значительным кормом. Нет никаких оснований предполагать, что девонские рыбы были менее благоразумными, чем современные.По словам Мелины Хейл из Чикагского университета, не все древние тропы были созданы ранними четвероногими, но также могли быть созданы родственниками четвероногих, которые использовали свои мясистые придатки для подобного передвижения на основе субстрата.
Исследования Дженнифер А. Клак и ее коллег показали, что самые ранние четвероногие животные, похожие на Acanthostega, были полностью водный инепригодный для жизни на суше. Это контрастирует с более ранним представлением о том, что сначала вторглась на сушу - либо в поисках добычи (как современные прыгуны ), либо для того, чтобы найти воду, когда пруд, в котором они жили, высох - а позже развились ноги, легкие и т. д.
К девона наземные растения стабилизировали пресноводные среды среды обитания, которые позволяют водно-болотным экосистемам развиваться, причем постоянно пищевые сети, открывшие новые возможности. Пресноводные среды обитания были не единственными местами, где можно было найти воду, наполненные органическими веществами и густой растительностью, у самой кромки воды. В этой среде также существуют болотистые места обитания, такие как мелкие водно-болотные угодья, прибрежные лагуны и большие солоноватоводные речные дельты, и есть много оснований предполагать, что именно в такой среде развивались четвероногие. Ранние ископаемые четвероногие были обнаружены вморских отложениях, потому что окаменелости примитивных четвероногих обычно разбросаны по всему миру, они, должно быть, распространились, следуя прибрежным линиям - они не могли жить только в пресной воде.
Один анализ, проведенный в Университете Орегона, предполагает отсутствие доказательств теории «сужающейся водяной скважины» - переходные окаменелости не связаны с доказательствами сужения луж или прудов - и указывает на то, что такие животные, вероятно, неили бы в в коротких переходахмежду истощенные водоемы. Вместо этой новой теории предполагается, что протолёгкие и прото-конечные полезные приспособления для преодоления окружающей среды во влажных лесных поймах.
Девонские четвероногие прошли через два основных узких места во время так называемого позднего периода. Девонское вымирание ; один в конце франского этапа, и один вдвое больше в следующем фаменского этапа. Эти события вымирания приводят к исчезновению примитивных четвероногих срыбоподобными чертами, таких как Ichthyostega, и их основных водных родственников. Когда четвероногие снова появляются в летописи окаменелостей после девонского вымирания, все взрослые формы полностью адаптированы к наземному существованию, более поздние вторично адаптированы к водному образу жизни.
общий предок всех нынешних гнатостомов (челюстно-позвоночные) в пресной воде, а позже мигрировал обратно в море. Чтобы справиться с такой болеевысокой соленостью морской воды, они разработали способность превращать азот отходы в безвредную мочевину, сохраняя ее в организме для дать крови же осмолярность, что и морская вода, не отравляя организм. Это система, которая в настоящее время используется у хрящевых рыб. рыбы с плавниками (Actinopterygii) позже вернулись в пресную воду и потеряли эту способность, в то время как рыбы с плавниками (Sarcopterygii) сохранилиее. Кровь из аммиака через жабры. Когда они, наконец, снова вернулись в море, они не применили свой старый трюк с превращением аммиака в мочевину, и вместо этого им пришлось развить солевые железы. Легкие рыбы делают то же самое, когда они живут в воде, производя аммиак, а не мочевину, но когда вода высыхает и они вынуждены зарываться в грязь, они переключаются на производство мочевины. Подобно хрящевым рыбам, латимерия может накапливать мочевину в своей крови, как иединственные известные земноводные, которые могут долгое время жить в соленой воде (жаба Bufo marinus и лягушка Rana cancrivora ). Это черты, которые они унаследовали от своих предков.
Использовали только аммиак, потребовалось бы позже его заново с нуля. Ни один из ныне живущих здесь рыб с лучевыми плавниками не смог этого сделать, поэтому маловероятно, что четвероногие тоже поступили бы так. Наземные животные, которые могут производитьтолько аммиак, должны будут постоянно пить, что делает жизнь на суше невозможной (существует несколько исключений, поскольку некоторые наземные мокрицы могут выделять азотистые отходы в виде газообразного аммиака). Вероятно, это также было проблемой вначале, когда четвероногие начали проводить время без воды, но в конечном итоге система мочевины стала полностью доминировать. Из-за этого маловероятно, что они появились в пресной воде (если только они сначала не мигрировали впресноводные среды обитания, а затем мигрировали на сушу вскоре после этого, они все еще сохраняли способность производить мочевину), хотя некоторые виды никогда не покидали воду или не возвращались в нее. могли, конечно, адаптироваться к пресноводным озерам и рекам.
Теперь ясно, что у общего предка костистых рыб (Osteichthyes) было примитивное дыхание воздухом легкое - позже эволюционировало в плавать мочевой пузырь у большинства актиноптеригов ( лучеплавниковых рыб). Это говорит о том, что кроссоптериги эволюционировали на теплых мелководьях, используя свое простое легкое, когда уровень кислорода в воде стал слишком низким.
Мясистые лопастные плавники, опирающиеся на кости, а не усиленные лучами плавники, по-видимому, были наследственной чертой всех костистых рыб (Osteichthyes ). Лопастоперые предки четвероногих развили их дальше, в то время как предки лучевых рыб (Actinopterygii ) развили свои плавники вдругом направлении. У наиболее примитивной группы актиноптеригов, бихиров, лобные плавники мясистые.
Были описаны девять родов девонских четвероногих, некоторые из которых известны в основном или полностью из материала нижней челюсти. Все, кроме одного, были с Лавразийского суперконтинента, который включал Европу, Северную Америку и Гренландию. Единственноеисключение - единственный гондванский род, Metaxygnathus, который был найден в Австралии.
. Было обнаружено первое девонское четвероногое животное, идентифицированное в Азии. из ископаемой челюстной кости, обнаруженной в 2002 году. Китайский четвероногий Sinostega pani был обнаружен среди окаменелых тропических растений и лопастоперых рыб в красном песчанике отложения Нинся-Хуэй автономного районасеверо-западного Китая. Это открытие существенно расширило географический ареал этих животных и подняло новые вопросы о мировом распространении и большом таксономическом разнообразии, которого они достигли за относительно короткое время.
 Самые старые следы четвероногих из Зачелми в связи с ключевыми девонскими тетраподоморфами окаменелостями
Самые старые следы четвероногих из Зачелми в связи с ключевыми девонскими тетраподоморфами окаменелостями  Eusthenopteron
Eusthenopteron  Panderichthys
Panderichthys  Tiktaalik
Tiktaalik  Acanthostega
Acanthostega  Tuchthyostega
Tuchthyostega  Hynerpetlerpeton
Hynerpetlerpeton 
 Crassigyrinus
Crassigyrinus  Diadectes
Diadectes Эти самые ранние четвероногие не были земными. Самые ранние подтвержденные земные формы известны из отложений начала карбона, примерно 20 миллионов лет спустя. Тем не менее, они могли проводить очень короткие периоды без воды и использовали бы свои ноги, чтобы проложить себе путь через грязь.
. Почему они вообще пошли на сушу, все еще обсуждается. Одной из причин могло быть то, что маленькие подростки, завершившие свою метаморфозу, имели всенеобходимое, чтобы использовать то, что предлагала земля. Уже приспособлены дышать воздухом и передвигаться по мелководью вблизи суши в качестве защиты (точно так же, как современные рыбы и земноводные часто проводят первую часть своей жизни в относительной безопасности мелководий, таких как мангровые леса), два очень разные ниши частично перекрывали друг друга, между ними находилась молодая молодь. Один из них был переполнен и опасен, в то время как другой был намного безопаснее именее многолюдным, предлагая меньшую конкуренцию за ресурсы. Наземная ниша также была гораздо более сложным местом для в первую очередь водных животных, но из-за того, как работают эволюция и давление отбора, те молодые особи, которые могли бы воспользоваться этим, будут вознаграждены. Как только они закрепятся на суше, благодаря их предварительной адаптации, благоприятные изменения в их потомках постепенно приведут к продолжению эволюции и диверсификации.
В это время множествобеспозвоночных, ползающих по суше и у воды, во влажной почве и мокрой подстилке, служило источником пищи. Некоторые были даже достаточно большими, чтобы поедать маленьких четвероногих, но земля была свободна от опасностей, типичных для воды.
Первоначально совершая лишь пробные набеги на сушу, четвероногие со временем адаптировались к земным условиям и проводили более длительные периоды вдали от воды. Также возможно, что взрослые особи начали проводить некотороевремя на суше (как предполагают модификации скелета ранних четвероногих, таких как Ichthyostega ), чтобы греться на солнце у кромки воды, в то время как в остальном они были в основном водными.
До 1990-х годов в летописи окаменелостей между четвероногими позднедевонскими и повторным появлением окаменелостей четвероногих в известном среднем карбоне <347 существовал разрыв в 30 миллионов лет>амфибия родословная. Онупоминался как «Промежуток Ромера », который теперь охватывает период примерно от 360 до 345 миллионов лет назад (девонско-каменноугольный переход и ранний период Миссисипи) после палеонтолога кто это узнал.
Во время «разрыва» у четвероногих развились позвоночник, а также конечности с пальцами и другие приспособления для земной жизни. Уши, черепа и позвоночные колонны тоже претерпели изменения. Число цифр на руках и ногахстало стандартизированным до пяти, поскольку линии с большим количеством цифр вымерли. Таким образом, те очень немногие окаменелости четвероногих, найденные в этом «промежутке», тем больше ценятся палеонтологами, потому что они документируют эти значительные изменения и разъясняют их историю.
Переход от водной рыбы с лопастными плавниками к земноводным, дышащим воздухом, был значительным и фундаментальным в истории эволюции позвоночных. Чтобы организм мог жить в нейтральнойводной среде с силой тяжести , затем колонизировать среду, которая требует, чтобы организм поддерживал весь его вес и обладал механизмом смягчения обезвоживания, требовались значительные адаптации или истощения в пределах всего план тела как по форме, так и по функциям. Эриопс, пример животного, сделавшего такую адаптацию, усовершенствовал многие черты его предков, рыб. Крепкие конечности поддерживали и перемещали его тело вне воды. Более толстый ипрочный хребет предохраняет его корпус от провисания под собственным весом. Кроме того, благодаря изменению формы рудиментарных костей челюстей рыбы начало развиваться рудиментарное среднее ухо, которое соединяется с внутренним ухом рыб, что позволяет Eryops усиливать и, таким образом, лучше воспринимать звуки, передаваемые по воздуху .
На визейском этапе (средний карбоновый период) первые четвероногие разошлись по крайней мере на три или четыре основные ветви. Некоторые из этих различных ветвей представляют собой предков всех живых существ. этраподы. Это означает, что общий предок всех современных четвероногих, вероятно, жил в каменноугольном периоде. Согласно узкому кладистическому определению тетрапод (также известного как корона-тетрапода), которое включает только потомков этого общего предка, четвероногие впервые появились в каменноугольном периоде. Узнаваемые ранние четвероногие (в широком смысле) являются представителями темноспондилов (например, Eryops ) лепоспондилов (например, Diplocaulus ), антракозавры, wh Они были родственниками и предками Амниоты и, возможно, бафетидов, которые, как полагают, связаны с темноспондилами и чей статус в качестве основной ветви пока не решен. В зависимости от того, каким авторитетным источникам следует следовать, современные земноводные (лягушки, саламандры и цецилии ), скорее всего, произошли от темноспондилов или лепоспондилов (или, возможно, от обоих, хотя это теперь позиция меньшинства).
Первый амниот (клад of vertebrates that today includes reptiles, mammals, and birds ) are known from the early part of the Late Carboniferous. By the Triassic, this group had already radiated into the earliest mammals, turtles, and crocodiles (lizards and birds appeared in the Jurassic, and snakes in the Cretaceous ). This contrasts sharply with the (possibly fourth) Carboniferous group, t he baphetids, which have left no extant surviving lineages.
Amphibians and reptiles were strongly affected by the Carboniferous rainforest collapse (CRC), an extinction event that occurred ~307 million years ago. The Carboniferous period has long been associated with thick, steamy swamps and humid rainforests. Since plants form the base of almost all of Earth's ecosystems, any changes in plant distribution have always affect до некоторой степени уменьшил животную жизнь. Внезапный коллапс жизненно важной экосистемы тропических лесов сильно повлиял на разнообразие и численность основных групп четвероногих, которые полагались на нее. CRC, который был частью одного из двух самых разрушительных исчезновений растений в истории Земли, был самоусиливающимся и очень быстрым изменением окружающей среды, в результате которого климат во всем мире в целом стал намного суше и прохладнее (хотя в настоящее время проводится много новой работы. чтобы лучше понять мелкие исторические
{kind=link}