| Амниоты | | Зауропсиды (включая динозавров)  | | | Синапсиды | | казеиды | Cotylorhynchus  | | | Eupelycosaurs | | Edaphosaurids | Edaphosaurus  | | | Sphenacodontians | | | | | | Perapsidsalc871>MСрок службы Это окно: -4500 - –-4000 - –-3500 - –-3000 - – -2500 - –-2000 - – -1500 - –-1000 - –-500 - –0 - Вода Одноклеточные. жизнь Фотосинтез Эукариоты Многоклеточная. жизнь Членистоногие Моллюсы Растения Динозавры Мящие Цветы Птицы Приматы ←Земля (−4540 )←Древнейшая вода ←Ранняя жизнь ←Самый ранний кислород ←Атмосферный кислород ←Кислородный кризис ←Половое размножение ←Древние растения ←Древние животные ← Кембрийский взрыв ←Тетрапода ←Древнейшие обезьяны P. h. a. n. e. r. o. z. o. i. c ....... P. r. o. t. e. r. o. z. o. i. c ... A. r. c. h. e. a. n H. a. d. e. a. n Понго ла Гу ронские Криогенные Андские Кару Четвертичные Ледниковые периоды ←Древнейшие грибы (миллионов лет назад ) первыми полностью наземными позвоночными были амниоты - их яйца имели внутренние мембраны, которые позволяли развивающемуся эмбриону дышать, но сохраняли воду. Это позволяет амниотам откладывать яйца на суше на суше, в то время как амфибии обычнонеобходимо откладывать яйца в воде ( некоторые амфибии, такие как обыкновенная суринамская жаба, развили другие способы обойти это ограничение). Первые амниоты, по-видимому, произошли в середине карбона от предков рептилиоморфов.
. В течение нескольких миллионов лет две важные линии амниот стали отличаться: предки синапсидов млекопитающих и сауропсиды, от которых произошли ящерицы, змеи, черепахи / черепахи, крокодилы,динозавры и птицы. Самые ранние изизвестных окаменелостей синапсидов и завропсид (таких как Archaeothyris и Hylonomus соответственно) датируются примерно 320–315 миллионами лет назад. Значительно раньше, чем первые ископаемые. 82> Синапсиды Исходная структура черепа синапсида имеет одно отверстие позади каждого, в довольно низком положении на этом черепе (внизу справа на изображении). Исходная структура черепа синапсида имеет одно отверстие позади каждого, в довольно низком положении на этом черепе (внизу справа на изображении). Синапсид черепа идентифицируются по характерному рисунку изотверстий позади каждого глаза, который служил следующим глазам: - делал череп легче без ущерба для прочности.
- экономия энергии за счет использования меньшего количества кости.
- , вероятно, обеспечивал точки прикрепления мускулов челюсти. Наличие прикрепления от челюсти позволяет мышцам стать длиннее и, следовательно, иметь сильное напряжение в широком диапазоне движений челюсти без растяжения или сокращения их оптимального диапазона.
Синапсид пеликозавры включал наземныхпозвоночных ранней перми, таких как 6 м (20 футов) длиной Cotylorhynchus hancocki. Среди других крупных пеликозавров были Dimetrodon grandis и Edaphosaurus cruciger. Therapsids Therapsids, произошедшие от пеликозавров в средней перми и занявшие их как доминирующие наземные позвоночные. Они отличаются от пеликозавровыми особенностями черепа и челюстей, включая более крупные височные отверстия и резцы, равные поразмеру.
Линиясидия, которая привела к млекопитающим прошли через ряд стадий, начиная с животных, очень похожих на своих предков-пеликозавров, и заканчивая другими, которые были легко приняты за млекопитающих: - соответствующее развитие костного вторичного небо. Большинство книг интерпретируют науку для эволюции высокой скорости метаболизма млекопитающих. Некоторые ученые отмечают, что некоторые современные эктотермы используют мясистое вторичное небо,чтобы отделить рот от дыхательных путей, и что костное небо обеспечивает поверхность, на которой язык может управлять пищей, облегчая жевание, а не дыхание. Интерпретация костного вторичного как вспомогательного средства при жевании также предполагает развитие более быстрого быстрого быстрого метаболизма, потому что это же сокращение размеров частиц, доставляемых в желудок, и, следовательно, может ускорить их переваривание. У млекопитающих, образованныхопределенными костями, различныелекарственные препараты имеют другие комбинации костей в нужных местах, чтобы функционировать как нёбо.
- зубная часть постепенно становится основной костью. нижняя челюсти.
- продвижение к вертикальной позе конечностей, которая поддерживает выносливость животных, избегая ограничений Карриера. Этот процесс был беспорядочным и очень медленным - например: у всех травоядных препаратов сохранились раскидистые конечности ( некоторые поздние возможности иметьполустоядные задние формы); Пермские плотоядные препараты имели раскидистые передние конечности, а у некоторых позднепермских также были полурасположенные задние конечности. Фактически, у современных монотрем все еще есть полуразвивающиеся конечности.
Генеалогическое древо терапии(упрощено; ниже возможности только те, которые имеют наибольшее отношение к эволюции млекопитающих) | |
Только дицинодонты, тероцефалы и тринодонты дожили доаса. БиармозухияБиармозухия были самыми примитивными и похожими на пеликозавровровыми терапиями. ДиноцефалыДиноцефалы ("ужасный головы"), включающие как плотоядных, так и травоядных. Они были большими; Антеозавр был до 6 м (20 футов)в длину. У некоторых хищников былиполуотстоящие задние конечности, но у всех диноцефалов они были очень примитивными терапиями; например, у них не было вторичного неба, а их челюсти были скорее «рептильными». Аномодонты Lystrosaurus, один из немногих родов дицинодонтов, переживших пермско-триасовое вымирание Lystrosaurus, один из немногих родов дицинодонтов, переживших пермско-триасовое вымирание аномальные зубы («аномальные зубы») были одними из самых успешных среди травоядных терапий - одна подгруппа, дицинодонты, дожила почти доконца триаса. Но аномодонты сильноотличались от современных травоядных млекопитающих, поскольку их единственные зубы были парой клыков в верхних челюсти, и принято считать, что у них были клювы, как у птиц или цератопсов. териодонтов<331 У териод онтов («звериные зубы») и их потомков челюстные суставы, в которых суставная кость нижней челюсти плотно сжимала очень маленькую квадратную кость черепа. Это позволяет получить гораздо более широкоезрение, и одна группа, хищные горгонопсии («лица горгон»), воспользовалась этим, чтобы развить «саблезубые». Увеличенный размер квадратной кости важен в развитии челюстного сустава и среднего уха млекопитающих.
У горгонопсийцев все еще были некоторые примитивные черты: отсутствие костного вторичного неба (но другие кости в нужных местах для выполнения тех же функций); раздвинутые передние конечности; задние конечности, которые могли работать как в раскинутых, так и ввертикальных позах. Но тероцефалы ( «звериные головы»), которые, по-видимому, возникли примерно в то же время, что и горгонопсии, обладали дополнительными чертами, похожими на млекопитающих, например их кости пальцев рук и ног имели такое же количество фаланг (сегментов), как и ранних млекопитающих (и такое же количество, как у приматов, включая людей). Цинодонты Концепция художника цинодонт триракодон в норе Концепция художника цинодонт триракодон в норе Цинодонт , группатериодонтов, возникшая также в конце перми,включает предков всех млекопитающих. Характерные черты цинодонтов, напоминающих млекопитающих, включая дальнейшее сокращение количества костей в нижних челюсти, вторичное костное небо, щечные зубы со сложным рисунком на коронках и мозгах, заполняющий эндокраниальную полость.
- найдены камерные норы, до 20 скелетов раннетриасового цинодонта трираходона ; Предполагается, что животные утонули в результате внезапногонаводнения . Обширные общие данные указывают на то,что эти данные могут быть к сложному социальному поведению. Триасовый захват Катастрофическое массовое вымирание в конце пермского периода, около 252 миллионов лет. назад погибли около 70 процентов наземных видов позвоночных и основных наземных растений.
В результате экосистемы и пищевые цепи рухнули, и на создание новых стабильных экосистем ушло около 30миллионов лет. Исчезновение горгонопсий, которые являютсядоминирующими хищниками поздней перми, перед конкурентами цинодонтов за доминирование в хищных нишах, малоизвестная группа сауропсид, архозавров, которые входят в состав предки крокодилов и динозавры.
Архозавры быстро стали доминирующими хищниками, и это явление часто называют «захватом триаса». Их успех мог быть связан с тем, что ранний триас был преимущественно засушливым, и поэтому превосходная сохранность водыархозаврами дала им решающее преимущество. Все известныеархозавры имеют кожу без желез и удаляют азотистые отходы в пасте мочевой кислоты, имеющей мало воды, в то время как цинодонты, вероятно, выделяли большую часть таких отходов в растворе мочевины, как млекопитающие сегодня; для растворения мочевины требуется значительное количество воды.
Однако эта теория была подвергнута сомнению, поскольку она подразумевает, что синапсиды имеют меньшиепреимущества в удержании воды, что сокращение синапсидов приводит кизменениям климата или разнообразию архозавров (ни один из них были протестированы) и тот факт, что обитающие в пустыне млекопитающих так же хорошо адаптированы в этом отделе, как и архозавры, некоторые цинодонты, такие как Trucidocynodon, были крупными хищниками.
Захват триаса, вероятно, был жизненно важный фактор в эволюции млекопитающих. Две группы, происходящие от ранних цинодонтов, добились успеха внишах, в которых конкуренция со стороны архозавров была минимальной:тритилодонты, которые были травоядными, и млекопитающие, большинство из которых великих ночной образ жизни насекомые (хотя некоторые из них, например Sinoconodon, были плотоядными животными, питавшимися добычей позвоночных, тогда как третьи были травоядными или всеядными ). В результате: - Тенденция терапевсида к дифференцировке зубов с точной окклюзией усилилась из-за необходимости удерживать пойманных членистоногих ираздавливать их экзоскелеты.
- Длина тела предков млекопитающих упавших ниже 50 мм (2 дюймов), достижения в области теплоизоляции и регулирование температуры стали бы необходимыми для ночного образа жизни.
- обострились слух и обоняние.
- Это ускоряло развитие среднего уха млекопитающих.
- Увеличение размера обонятельных долей мозга увеличило массу в процентах от общеймассы тела. Мозговая ткань требует непропорционально много энергии. Потребностьв увеличении количества пищи для поддержки увеличенного мозга давление на улучшение теплоизоляции, регулирования температуры и кормления.
- Вероятно, в результате ночного образа жизни млекопитающих потеряли две из четырех колбочек опсинов, фоторецепторы в сетчатке, присутствующие в истории самых ранних амниот. Парадоксально, но это могло улучшить их способность различать цвета при тускломсвете.
Это отступление к ночной роли узким местом в ночное время и,как считается, объясняет многие особенности млекопитающих. От цинодонтов до верхних млекопитающих Летопись окаменелостейМезозойские синапсиды, которые эволюционировали до состояния челюстного сустава, состоящего из зубных и чешуйчатых костей, сохранились в нескольких хороших окаменелостях, по существу потому что они были в основном хорошие, чем крысы: - Они в основном былиограничены средой, которая с меньшей вероятностью дает окаменелости. Поймы, как лучшая земная среда для окаменения, дают мало окаменелостей млекопитающих, потому что в них преобладают средние и крупные животные, которые могут конкурировать с архозаврами в диапазоне от средних до крупных. Следы из раннего мела Анголы показывают наличие млекопитающих размером с енота 118 миллионов лет назад.
- Их хрупкие кости были уязвимы для разрушения, прежде чем онимогли быть окаменелы - падальщиками (включая грибы и бактериями ) инаступлением на них.
- Маленькие окаменелости труднее разрушительного и более уязвимы для разрушения в результате выветривания и других природных ресурсов, прежде чем они будут обнаружены.
Однако за последние 50 лет количество ископаемых мезозойских млекопитающих значительно увеличилось; только 116 родов были известны в 1979 году, например, но около 310 в 2007 году, с повышением качества, такчто «по крайней мере 18 мезозойских млекопитающих представлены почти полными скелетами». Млекопитающие или формы млекопитающихНекоторые авторы ограничивают термин «млекопитающее» группой крон млекопитающих, группой, состоящей из самых недавних общих предков монотрем, сумчатых, и плацентарные вместе со всеми потомками этого предка. В влиятельной статье 1988 года Тимоти Роу отстаивал это ограничение, утверждая, что «происхождение...обеспечивает единственное средство для правильного определения таксонов» и, в частности, чтоотклонение однообразных от животных, более тесно связанных с сумчатыми и плацентарями » представляет центральный интерес для любого исследования млекопитающих в целом ». Чтобы учесть некоторые родственные таксоны, не входящие в группу кроны, он определил Mammaliaformes как включающий «последнего общего предка Morganucodontidae и Mammalia [как он определил последний термин] и всех их потомков».Помимо Morganucodontidae, новый таксон включает Docodonta и Kuehneotheriidae. Хотя харамиидыотносились к млекопитающим с 1860-х годов, Роу исключил их из группы млекопитающих как выходящих за рамки своего определения, поместив их в более крупную кладу, млекопитающих.
, чтобы избежать недоразумений, некоторые авторы приняли эту терминологию., что они это сделали. Однако большинство палеонтологов все еще считают, что животные с зубно-чешуйчатым челюстным суставом и молярами,характерными для современных млекопитающих, формально должны быть членами Mammalia.
Гдедвусмысленность в термине «млекопитающее» может сбивать с толку, в этой статье используются «млекопитающие» и «кроны млекопитающих». Семейное древо - цинодонты к млекопитающим группы кроны(на основе Cynodontia: Dendrogram - Palaeos ) | |
| |
| |
| |
 Morganucodontidae и другие переходные формы имели оба типа челюстных суставов: зубной - чешуйчатый (передний) и суставной - квадратной (сзади). Morganucodontidae и другие переходные формы имели оба типа челюстных суставов: зубной - чешуйчатый (передний) и суставной - квадратной (сзади). MorganucodontidaeMorganucodontidae впервые появились в конце триаса, около 205миллионов лет назад.. Они являются прекрасным примером переходных окаменелостей, поскольку имеют какзубно-чешуйчатые, так и суставно-квадратные челюстные суставы. Они также были одними из первых обнаруженных и наиболее тщательно изученных млекопитающих. s за пределами коронной группы млекопитающих, поскольку было обнаружено необычно большое количество окаменелостей морганукодонтов. Докодонты Реконструкция Касторокауды. Обратите внимание на мех иприспособления для плавания (широкий плоский хвост; перепончатые лапы) и рытья рук (крепкие конечности икогти). Реконструкция Касторокауды. Обратите внимание на мех иприспособления для плавания (широкий плоский хвост; перепончатые лапы) и рытья рук (крепкие конечности икогти). Докодонты, среди наиболее распространенных юрских млекопитающих, известны изощренностью своих коренных зубов.. Считается, что у них были общие полуводные наклонности, например, рыбоядные Castorocauda («бобровый хвост»), которые жили в середине юрского периода около 164 миллионов лет назад и были Впервые обнаружен в 2004 г. и описан в 2006 г. и являетсянаиболее понятным примером. Castorocauda не относилось к млекопитающим коронной группы, но оночрезвычайно важно для изучения эволюции млекопитающих, потому что первой находкой был почти полный скелет (настоящая роскошь в палеонтологии), и он ломает стереотип о «маленьком ночном насекомоядном»: - Он был заметно больше, чем большинство окаменелостей мезозойских млекопитающих - около 17 дюймов (43 см) от носа до кончика 5-дюймового (130 мм) хвоста и, возможно, весил 500–800 г (18–28 унций).
- Это самые ранние, абсолютно достоверные свидетельства наличия волос и меха. Ранеесамым ранним из них был Eomaia, млекопитающее коронной группы, появившееся около 125 миллионов лет назад.
- У него были водные приспособления, включая сплющенные кости копыта и остатки мягких тканей между пальцами задних ног, что позволяет предположить что они были перепончатыми. Ранее самые ранние известные полуводные формы млекопитающих были из эоцена, примерно 110миллионов лет спустя.
- Мощные передние конечности Касторокауды выглядят приспособленнымидля рытья. Эта особенность и шпоры на лодыжках делают его похожим на утконоса, который также плавает и копает.
- Его зубы выглядят приспособленными для поедания рыбы: первые два коренных зуба имели бугорки в прямом ряду., что сделало их более удобными для захвата и нарезки, чем для измельчения; и эти коренные зубы изогнуты назад, чтобы помочь хватать скользкую добычу.
HadrocodiumНа приведенном выше генеалогическом древе Hadrocodium изображен как«тетя» кроновых млекопитающих. Эта форма млекопитающего, датируемая примерно 195 млн. Лет назад в самом раннем юрском периоде, демонстрирует некоторые важные особенности: - Челюстной сустав состоит только из чешуйчатых и зубных костей, а челюсть не содержит более мелких костей в задней части зубной кости, в отличие от
- В терапсидах и ранних формах млекопитающих барабанная перепонка могла растягиваться над впадиной в задней части нижней челюсти. Но у Hadrocodiumне было такого впадины, что позволяет предположить, что его ухо было частью черепа, как у млекопитающих коронной группы - и, следовательно, бывшие суставные и квадратные переместился в среднее ухо и превратился в молоток и наковальню. С другой стороны, у зубной кости есть «бухта» сзади, которой нет у млекопитающих. Это говорит о том, чтозубная кость Hadrocodium сохранила ту же форму, которую имела бы, если бы суставная и квадратная частиоставались частью челюстного сустава, и, следовательно, Hadrocodium или его очень близкий предок, возможно, были первыми, у кого было полностью среднее ухо млекопитающих.
- Терапсиды и более ранние формы млекопитающих имели челюстные суставы очень далеко в черепе, отчасти потому, что ухо находилось в заднем конце челюсти, но также должно было быть близко к мозгу. Такое расположениеограничивало размер мозговой коробки, потому что заставляло мышцы челюсти бегать по ней. Мозговая оболочкаи челюсти Hadrocodium больше не были связаны друг с другом из-за необходимости поддерживать ухо, а его челюстной сустав был выдвинут вперед. У его потомков или у потомков животных с аналогичным расположением корпус мозга мог свободно расширяться, не ограничиваясь челюстью, и челюсть могла свободно меняться, не ограничиваясь необходимостью держать ухо рядом с мозгом - другими словами теперьстало возможным для млекопитающих как развить крупный мозг, так и приспособить свои челюсти и зубы так,чтобы они были полностью специализированы для приема пищи.
Древнейшие коронные млекопитающие коронная группа млекопитающие, иногда так называемые «настоящие млекопитающие» - это современные млекопитающие и их родственники до их последнего общего предка. Поскольку в этой группе есть живые члены, ДНК анализ может быть применен в попытке объяснитьэволюцию особенностей, которые не проявляются в окаменелостях. Это стремление часто включает молекулярную филогенетику, метод, который стал популярным с середины 1980-х годов. Родословная древних млекопитающих кроныКладограмма по З.-Х Луо. († обозначает вымершие группы) ЦветовоезрениеРанние амниоты имели четыре опсина в колбочках сетчатки, которые использовались для различения цвета: один чувствителен к красному, один - к зеленому и два - к разным оттенкам синего. Зеленый опсин не был унаследован никакими коронными млекопитающими, но все нормальные особи унаследовали красный. Таким образом, у ранних кроновых млекопитающих было три колбочковых опсина,красный и оба голубых. Все их нынешние потомки потеряли один из чувствительных к синему опсинов, но не всегдаодин и тот же: монотремы сохраняют один чувствительный к синему опсин, в то время как сумчатые и плацентарные животные сохраняют другой (кроме китообразных, которые позже потеряли второй опсин. синий опсин). У некоторых плацентарных и сумчатых животных, включая высших приматов, впоследствии развились опсины, чувствительные к зеленому; поэтому, как и у ранних коронныхмлекопитающих, их зрение трехцветное. Australosphenidae и AusktribosphenidaeAusktribosphenidae -название группы, которое было дано некоторым довольно загадочным находкам: - , по-видимому, трибосфенические коренные зубы, тип зубов, который иначе известен только у плацентарных и сумчатых животных.
- происходят из отложений среднего мелового периода в Австралии, но Австралия была связана только с Антарктидой, и плаценты возникли в Северномполушарии и были приурочены к нему до тех пор, пока дрейф континентов не сформировал сухопутные связи изСеверной Америки в Южную Америку, из Азии в Африку и из Азии в Индию (карта позднего мела здесь показывает, как разделены южные континенты).
- представлены только зубами и фрагментами челюстей, что не очень полезно.
Australosphenida - это группа, которая была определена для включения Ausktribosphenidae и monotremes. Asfaltomylos (mid- to lateJurassic, from Patagonia ) has been interpreted as a basal australosphenid (animal that has features sh встречаются как с Ausktribosphenidae, так и с монотремами; не имеет признаков, присущих Ausktribosphenidae или monotremes; также отсутствуют черты, отсутствующие у Ausktribosphenidae и monotremes), и это свидетельствует о том, что австралофениды были широко распространены на всей территории Гондваны (старый суперконтинент Южного полушария).
Недавний анализ Teinolophos, который жил где-то между 121 и 112,5 миллионами лет назад, предполагает, что это была «группа короны» (развитая и относительная у специализированный) monotreme. Это было воспринято как доказательство того, что базальные (наиболее примитивные) монотремы должны были появиться значительно раньше, но это оспаривается (см. Следующий раздел). Исследование также показало, что некоторые предполагаемые австралосфениды также были монотремами «коронной группы» (например, Steropodon ) и что другие предполагаемые австралофениды (например, Ausktribosphenos, Bishops, Ambondro, Asfaltomylos) более тесно связаны и, возможно, являются членами группы Терианские млекопитающие (группа, включающая udes marsupials and placentals, see below). MonotremesTeinolophos, from Australia, is the earliest known monotreme. A 2007 study (published 2008) suggests that it was not a basal (primitive, ancestral) monotreme but a full-fledged platypus, and therefore th у утконоса и ехидны родословные расходились значительно раньше. Однако более недавнее исследование (2009 г.) показало, что, хотя теинолофос был типом утконоса, он также был базальной монотремой, и данные свидетельствуют о том, что ехидны отделились от линии утконоса еще в 19-48 мес, когда они сделали свое переход от полуводного к наземному образу жизни.
Однородные имеют некоторые особенности, которые могут быть унаследованы от предков цинодонтов : - как ящерицы and birds, they use the same orifice to urinate, defecate and reproduce ("monotreme" means "one hole").
- they lay eggs that are leathery and uncalcified, like those of lizards, turtles andcrocodilians.
Unlike other mammals, female monotremes do not have nipples and feed their young by "sweating" milk from patches on their bellies.
These features are not visible in fossils, and the main characteristics from paleontologists' point of view compared to m у арсупиалов и плацентарных млекопитающих - около 32 ° C (90 ° F). Филогенетический брекетинг предполагает, что температура тела ранних млекопитающих коронной группы была не ниже, чем у современных монотрем. Имеются цитологические свидетельства того, что низкий метаболизм монотремов является вторично развитой чертой. Дыхательные носовые раковиныУ современных млекопитающих есть дыхательные раковины, извитые структуры тонкой кости в носовой полости полость. Они выстланы слизистыми оболочками, которые warm and moisten inhaled air and extract heat and moisture от выдыхаемого воздуха. Животное с носовыми раковинами может поддерживать высокую частоту дыхания без опасности пересыхания легких и, следовательно,может иметь быстрый метаболизм. К сожалению, эти кости очень хрупкие и поэтому еще не были обнаружены в окаменелостях. Но рудиментарные гребни, подобные тем, которые поддерживают носовые раковины, были обнаружены у продвинутых триасовых цинодонтов, таких как Thrinaxodon и Diademodon, что предполагает, что они могли иметь довольно высокий скорость метаболизма. Костное вторичное небоУ млекопитающих есть вторичное костное небо, которое отделяетдыхательные пути от рта, что позволяет им есть и дышать одновременно. Вторичное костное нёбо было обнаружено у более продвинутых цинодонтов и использовалось в качестве доказательства высокого уровня метаболизма. Но у некоторых хладнокровных позвоночных есть вторичное костное нёбо (крокодилы инекоторые ящерицы), тогда как у теплокровных птиц его нет. Диафрагмамускулистая диафрагма помогает млекопитающим дышать, особенно во время физических нагрузок. Чтобы диафрагмаработала, ребра не должны ограничивать живот, так что расширение грудной клетки может быть компенсировано уменьшением объема живота и наоборот. Диафрагмы известны у казеидных пеликозавров, что указывает на их раннее происхождение в синапсидах, хотя они все еще были довольнонеэффективными и, вероятно, требовали поддержки со стороны других групп мышц и движений конечностей.
У продвинутых цинодонтов есть очень млекопитающие. грудные клетки с сильно сокращенными поясничнымиребрами. Это говорит о том, что у этих животных были более развитые диафрагмы, они были способны к напряженной деятельности в течение довольно длительных периодов времени и, следовательно, имели высокий уровень метаболизма. С другой стороны, эти грудные клетки, подобные млекопитающим,могли развиться, чтобы повысить подвижность. Однако движение даже продвинутых терапсидов было «подобно тачке», когда задние конечности обеспечивали весь толчок, в то время как передние конечности только управлялиживотным, иными словами, продвинутые терапсиды были не такими проворными, как современные млекопитающие или ранние динозавры. Таким образом, идея о том, что основная функция грудных клеток, подобных млекопитающим, заключалась в повышении подвижности, сомнительна. ПозаконечностейУ терапсидов были раздвинутые передние конечности и полустоячие задние конечности. Это говорит о том, что ограничение Карриера сделало бы для них довольно трудным двигаться идышать одновременно, но не так сложно, как для таких животных, как ящерицы, у которых конечности полностью растянуты. Следовательно, усовершенствованные терапсиды могли быть значительно менее активными, чем современные млекопитающие того же размера, и поэтому, возможно, имели болеемедленный метаболизм в целом или же были брадиметаболическими (более низкий метаболизм в состоянии покоя). МозгМлекопитающие отличаются большим размером мозга по сравнению с размером телапо сравнению с другими группами животных. Недавние открытия предполагают, что первая область мозга, которая расширилась, была связана с запахом. Ученые просканировали черепа ранних видов млекопитающих, датируемых 190–200 миллионами лет назад, и сравнили формы корпуса мозга с болееранними видами, существовавшими до млекопитающих; они обнаружили, что область мозга, связанная с обонянием, увеличилась первой. Это изменение могло вызвать этим ранним млекопитающим охотиться на насекомых ночью, когдадинозавры не были активны. См. Также Ссылки Дополнительная литература Внешние ссылки - The Cynodontia охватывает несколько видов эволюции цинодонтов в млекопитающих, с большим количеством ссылок.
- Мучающие, обсуждение BBC Radio 4 с Ричардом Корфилдом, Стивом Джонсом и Джейн Фрэнсис (в наше время, 13 октября 2005 г.)
Последняя правка сделана 2021-05-19 09:15:26
Содержание доступно по лицензии CC BY-SA 3.0 (если не указано иное).
Обратная связь: support@alphapedia.ru

|

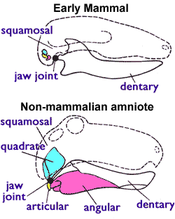 Рис. 1: У млекопитающих, квадратная и суставная кости маленькие и являются частью среднего уха ; нижняя челюсть состоит только из зубной кости.
Рис. 1: У млекопитающих, квадратная и суставная кости маленькие и являются частью среднего уха ; нижняя челюсть состоит только из зубной кости.  Исходная структура черепа синапсида имеет одно отверстие
Исходная структура черепа синапсида имеет одно отверстие 
 Концепция художника цинодонт
Концепция художника цинодонт 
 Реконструкция
Реконструкция