
Войти
| Евриптерид. Временной диапазон: Дарривилиан - Поздняя пермь, 467,3–251,9 Ma до Ꞓ O S D C P T J K Pg N Возможная тремадокская запись | |
|---|---|
 | |
| Ископаемый образец Eurypterus remipes, хранящийся в Государственном музее Естественная история Карлсруэ в Карлсруэ, Германия. | |
| Научная классификация | |
| Домен: | Эукариота |
| Царство: | Animalia |
| Тип: | Arthropoda |
| Subphylum: | Chelicerata |
| Clade: | Sclerophorata |
| Заказ: | †Eurypterida. Burmeister, 1843 |
| Suborders | |
| |
| Синонимы | |
Эвриптериды, часто неофициально называемые морскими скорпионами, представляют собой группу вымерших членистоногих, которые образуют отряд Эвриптерида . Самые ранние известные эвриптериды относятся к дарривильскому этапу ордовикского периода 467,3 миллиона лет назад. Группа, вероятно, впервые появилась либо в раннем ордовике, либо в позднем кембрии. Eurypterida насчитывает около 250 видов и представляет собой наиболее разнообразный палеозойский хелицератный отряд. После своего появления в ордовике, эвриптериды стали основными компонентами морской фауны в течение силурия, из которого было описано большинство видов эвриптерид. Силурийский род Eurypterus составляет более 90% всех известных экземпляров эвриптерид. Хотя группа продолжала диверсифицироваться в течение последующего девонского периода, эвриптериды сильно пострадали от позднедевонского вымирания. Они уменьшились в численности и разнообразии, пока не вымерли во время пермско-триасового вымирания (или когда-то незадолго до этого) 251,9 миллиона лет назад.
Хотя в народе их называли «морскими скорпионами», только самые ранние эвриптериды были морскими ; многие более поздние формы обитали в солоноватой или пресной воде, и они не были настоящими скорпионами. Некоторые исследования предполагают наличие двойной дыхательной системы, что позволяло бы в течение коротких периодов времени находиться в земной среде. Название Eurypterida происходит от древнегреческих слов εὐρύς (eurús), означающих «широкий» или «широкий», и πτερόν (pteron), что означает «крыло», относящихся к паре широких плавательных придатков, присутствующих в многие участники группы.
Эвриптериды включают в себя самых крупных известных из когда-либо существовавших членистоногих. Самый крупный, Jaekelopterus, достигал 2,5 метров в длину. Евриптериды не были одинаково крупными, и большинство видов были менее 20 сантиметров (8 дюймов) в длину; самая маленькая эвриптерида, Alkenopterus, была всего 2,03 сантиметра (0,80 дюйма) в длину. Окаменелости эвриптерид были обнаружены со всех континентов. Большинство окаменелостей происходит из окаменелостей в Северной Америке и Европе, потому что группа жила в основном в водах вокруг и внутри древнего суперконтинента Еврамерика. Лишь горстка групп эвриптерид распространилась за пределы Еврамерики, а несколько родов, таких как Adelophthalmus и Pterygotus, достигли космополитического распространения с ископаемыми останками, обнаруженными по всему миру..
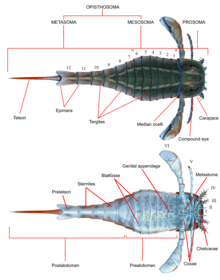 Восстановление Eurypterus с частями тела, помеченными
Восстановление Eurypterus с частями тела, помеченными Как и все другие членистоногие, эвриптериды обладали сегментированными телами и сочлененными придатками (конечностями), покрытыми кутикулой, состоящей из белков и хитина. Как и в других хелицератах, тело было разделено на две tagmata (секции); лобная просома (голова) и задняя опистосома (брюшная полость). Просома была покрыта панцирем (иногда называемым «просомальным щитом»), на котором были составные глаза и глазки (простые органы чувств, похожие на глаза).
Просома также несла шесть пар придатков, которые обычно называют парами придатков с I по VI. Первая пара придатков, единственная пара, помещенная перед ртом, называется хелицерами (гомологична клыкам пауков). У них были маленькие клешни, которыми они манипулировали фрагментами пищи и засовывали их в рот. В одной линии, Pterygotidae, хелицеры были большими и длинными, с сильными, хорошо развитыми зубами на специализированных челах (когтях). Последующие пары придатков, номера от II до VI, имели гнатобазы (или «зубные пластины») на тазиках (сегментах конечностей), используемых для питания. Эти придатки обычно представляли собой ходячие ноги, которые имели цилиндрическую форму и у некоторых видов были покрыты шипами. В большинстве линий конечности имели тенденцию становиться больше по мере удаления от себя. В подотряде Eurypterina , большем из двух подотрядов эвриптерид, шестая пара придатков также была преобразована в плавательное весло, чтобы помочь в перемещении по водной среде.
Опистосома состояла из 12 сегментов, а тельсон - самый задний сегмент, который у большинства видов имел форму лопатки. В некоторых линиях, в частности, у Pterygotioidea, Hibbertopteridae и Mycteroptidae, тельсон был сплющен и, возможно, использовался в качестве руля во время плавания. У некоторых родов суперсемейства Carcinosomatoidea, особенно Eusarcana, был тельсон, аналогичный таковому у современных скорпионов, и они могли использовать его для инъекции яд. Тазики шестой пары придатков были перекрыты пластиной, которая упоминается как метастома, первоначально полученная из полного сегмента экзоскелета. Сама опистосома может быть разделена на «мезосому » (включающая сегменты с 1 по 6) и «метасому » (включающая сегменты с 7 по 12) или на «преабдомен» (обычно включая сегменты с 1 по 7) и «постабдомен» (обычно содержащий сегменты с 8 по 12).
Нижняя сторона опистосомы была покрыта структурами, возникшими из модифицированных опистосомных придатков. Повсюду в опистосоме эти структуры образовывали пластинчатые структуры, называемые blatfüsse (немецкий для «листопада»). Они создали жаберную камеру (жаберный тракт) между предшествующим блатфюссом и вентральной поверхностью самой опистосомы, которая содержала органы дыхания. Со второго по шестой опистосомные сегменты также содержали овальные или треугольные органы, которые были интерпретированы как органы, способствующие дыханию. Эти органы, называемые kiemenplatten, или «жаберные пути», потенциально помогали эвриптеридам дышать воздухом над водой, в то время как blatfüssen, подобно органам современных подковообразных крабов, мог бы покрывать части, которые служат для подводное дыхание.
Придатки опистосомных сегментов 1 и 2 (седьмой и восьмой сегменты в целом) были слиты в структуру, названную генитальной крышкой, занимающей большую часть нижней стороны опистосомного сегмента 2. Около переднего отдела край этой структуры, генитальный придаток (также называемый зипфелем или средним брюшным придатком) выступал. Этот придаток, часто сохраняющийся очень заметно, последовательно интерпретируется как часть репродуктивной системы и встречается в двух признанных типах, которые, как предполагается, соответствуют мужчине и женщине.
 Сравнение размеров шести самых крупных видов эвриптерид из родов Jaekelopterus, Carcinosoma, Acutiramus (два вида), Pterygotus и Pentecopterus
Сравнение размеров шести самых крупных видов эвриптерид из родов Jaekelopterus, Carcinosoma, Acutiramus (два вида), Pterygotus и Pentecopterus Евриптериды сильно различались по размеру в зависимости от таких факторов, как образ жизни, среда обитания и таксономическая близость. Размеры около 100 сантиметров (3,3 фута) обычны для большинства групп эвриптерид. Самая маленькая эвриптерида, Alkenopterus burglahrensis, имела длину всего 2,03 см (0,80 дюйма).
Самая большая эвриптерида и самое крупное из известных членистоногих, когда-либо существовавших, это Jaekelopterus rhenaniae. Хелицера из эмсской формации Клерф в Виллверате, Германия имела длину 36,4 сантиметра (14,3 дюйма), но не имела четверти длины, что позволяет предположить, что полная хелицера была 45,5 сантиметра (17,9 дюйма) в длину. Если пропорции между длиной тела и хелицерами совпадают с пропорциями его ближайших родственников, где соотношение между размером когтей и длиной тела относительно стабильно, образец Jaekelopterus, который обладал рассматриваемой хелицерой, имел бы размеры от 233 до 259 сантиметров (от 7,64 до 8,50). футов), в среднем 2,5 метра (8,2 фута) в длину. Если хелицеры вытянуты, к этой длине будет добавлен еще один метр (3,28 фута). Эта оценка превышает максимальный размер тела всех других известных гигантских членистоногих почти на полметра (1,64 фута), даже если не учитывать расширенные хелицеры. Две другие эвриптериды также достигли длины 2,5 метра; Erettopterus grandis (близкий родственник Jaekelopterus) и Hibbertopterus wittebergensis, но E. grandis очень фрагментарен, и оценка размеров H. wittenbergensis основана на свидетельствах следов, а не на останках ископаемых.
Семейство Jaekelopterus, Pterygotidae, известно несколькими необычно крупными видами. Оба Acutiramus, самый крупный представитель которого A. bohemicus имел рост 2,1 метра (6,9 футов), и Pterygotus, у которого самый большой вид P. grandidentatus имел размеры 1,75 метра (5,7 футов), были гигантскими. Было предложено несколько различных факторов, способствующих большому размеру птеригид, включая ухаживание, хищничество и конкуренцию за ресурсы окружающей среды.
Гигантские эвриптериды не ограничивались семейством Pterygotidae. Изолированная ископаемая метастома длиной 12,7 см (5,0 дюйма) карциносоматоидной эвриптериды Carcinosoma punctatum указывает на то, что при жизни животное достигло бы в длину 2,2 метра (7,2 фута), конкурируя по размеру с птериготидом. Другим гигантом был Pentecopterus decorahensis, примитивный карциносоматоид, который, по оценкам, достиг в длину 1,7 метра (5,6 фута).
Типичным для крупных эвриптерид является легкое телосложение. Такие факторы, как передвижение, затраты энергии при линьке и дыхании, а также фактические физические свойства экзоскелета ограничивают размер, которого могут достигать членистоногие. Легкая конструкция значительно снижает влияние этих факторов. Pterygotids были особенно легкими, при этом большинство окаменелых крупных сегментов тела оставались тонкими и неминерализованными. Легкие приспособления присутствуют и у других гигантских палеозойских членистоногих, таких как гигантская многоножка Arthropleura, и, возможно, имеют жизненно важное значение для эволюции гигантских размеров у членистоногих.
В дополнение к легкому гиганту. eurypterids, некоторые глубокотелые формы семейства Hibbertopteridae также были очень крупными. Панцирь из карбона Шотландии, относящийся к виду Hibbertoperus scouleri, имеет ширину 65 см (26 дюймов). Поскольку Hibbertopterus был очень широким по сравнению со своей длиной, рассматриваемое животное могло иметь длину чуть менее 2 метров (6,6 футов). Этот гигантский Hibbertopterus, более прочный, чем птеригид, мог бы соперничать с самыми крупными птеригитами по весу, если бы не превосходил их, и, как таковой, был бы одним из самых тяжелых членистоногих.
 Иллюстрация подводного полета в Eurypterus, в котором формы лопастей и их движения в воде достаточно для создания подъемной силы. Этот тип передвижения ограничивался эвриптерином (с плавательными веслами), эвриптеридами.
Иллюстрация подводного полета в Eurypterus, в котором формы лопастей и их движения в воде достаточно для создания подъемной силы. Этот тип передвижения ограничивался эвриптерином (с плавательными веслами), эвриптеридами.  Иллюстрация Hibbertopterus, большой стилонурин (без плавательных лопастей) эвриптерид
Иллюстрация Hibbertopterus, большой стилонурин (без плавательных лопастей) эвриптерид Два подотряда эвриптерид, Eurypterina и Stylonurina, разделены в основном морфологией их последней пары придатков. У Stylonurina этот отросток принимает форму длинной и тонкой ходовой ноги, тогда как у Eurypterina нога видоизменена и расширена в плавательную лопатку. Если не считать плавательного весла, ноги многих эвриптеринов были слишком малы, чтобы делать что-то большее, чем позволять им ползать по морскому дну. Напротив, у некоторых сталонуринов были удлиненные и мощные ноги, которые могли позволить им ходить по суше (аналогично современным крабам ).
A ископаемым следам, обнаруженным в ископаемых отложениях каменноугольного возраста в Шотландии в 2005 году. был отнесен к стейлонуриновой эвриптериде Hibbertopterus из-за подходящего размера (длина гусеницы оценивается примерно в 1,6 метра (5,2 фута)) и предполагаемой анатомии ноги. Это самая большая наземная тропа длиной 6 метров (20 футов) и в среднем 95 см (3,12 фута) в ширину - сделанный членистоногим, обнаруженным на данный момент. Это первая запись эвриптерид, передвигающихся по суше. Путь пути свидетельствует о том, что некоторые эвриптериды могли выжить в земной среде, по крайней мере, в течение коротких периодов времени., и раскрывает информацию о походке стилонуринов. У Hibbertopterus, как и у большинства эвриптерид, пары придатков различаются по размеру (это называется гетероподвижным состоянием конечностей). Эти пары разного размера будут двигались синхронно, а короткий шаг указывает на то, что Hibbertopterus ползет с исключительно медленной скоростью, по крайней мере, на суше. Большого тельсона волочили по земле, оставляя за животным большую центральную бороздку. Наклоны дорожек через случайные интервалы предполагают, что движение было резким. Походка меньших по размеру стейлонуринов, таких как Parastylonurus, вероятно, была более быстрой и точной.
Функциональность плавательных лопастей из эвриптерина варьировалась от группы к группе. У Eurypteroidea весла по форме были похожи на весла. Состояние суставов их придатков гарантировало, что их лопасти можно было перемещать только в почти горизонтальной плоскости, а не вверх или вниз. Некоторые другие группы, такие как Pterygotioidea, не обладали этим состоянием и, вероятно, могли плавать быстрее. Принято считать, что большинство эвриптерин использовали гребной тип движения, подобный крабам и водяным жукам. Более крупные люди могли летать под водой (или подводный полет ), при котором движения и формы лопастей достаточно для создания подъемной силы, подобно плаванию в море. черепахи и морские львы. Этот тип движения имеет относительно более низкую скорость ускорения, чем гребной тип, особенно потому, что у взрослых особей весла пропорционально меньше, чем у молодых. Однако, поскольку большие размеры взрослых означают более высокий коэффициент лобового сопротивления, использование этого типа движителя более энергоэффективно.
 голотип из Palmichnium kosinkiorum, содержащий самые большие известные следы эвриптерид.
голотип из Palmichnium kosinkiorum, содержащий самые большие известные следы эвриптерид. Некоторые эвриптерины, такие как Mixopterus (как следует из приписываемых ископаемых следов), не обязательно были хорошими пловцами. Скорее всего, он держался в основном на дне, используя свои плавательные весла для случайных всплесков движений по вертикали, с четвертой и пятой парами придатков, расположенными назад, чтобы производить незначительное движение вперед. Во время ходьбы он, вероятно, использовал походку, как у большинства современных насекомых. Вес его длинного живота уравновешивался бы двумя тяжелыми и специализированными лобными придатками, а центр тяжести можно было бы регулировать, поднимая и располагая хвост.
Сохранившиеся ископаемые следы эвриптерид имеют тенденцию быть большими и неоднородными и часто имеют связанный след тельсона по средней линии (как в случае с шотландским следом Hibbertopterus). Такие тропы были обнаружены на всех континентах, кроме Южной Америки. В некоторых местах, где ископаемые останки эвриптерид в остальном редки, например, в Южной Африке и на остальной территории бывшего суперконтинента Гондвана, открытия тропинок предшествуют и превосходят по численности ископаемые останки тел эвриптерид. Следы эвриптерид были отнесены к нескольким ихногенерам, в первую очередь к Palmichnium (определенному как серия из четырех следов, часто с ассоциированной меткой перетаскивания в средней линии), где голотип ихновидов P. kosinkiorum сохраняет самые большие из известных на сегодняшний день следов эвриптерид, каждый из которых имеет диаметр около 7,6 см (3,0 дюйма). Другие eurypterid ichnogenera включают Merostomichnites (хотя вполне вероятно, что многие образцы действительно представляют собой следы ракообразных) и Arcuites (который сохраняет бороздки, образованные плавательными придатками).
 Предполагаемые «жаберные тракты» эвриптерид сравнивались с дыханием воздухом, присутствующим в задних ногах современных равноногих, таких как Oniscus (на фото)
Предполагаемые «жаберные тракты» эвриптерид сравнивались с дыханием воздухом, присутствующим в задних ногах современных равноногих, таких как Oniscus (на фото) У эвриптерид органы дыхания располагались на вентральной стенке тела (нижняя сторона опистосомы). Blatfüsse, произошедший от опистосомных придатков, покрыл нижнюю часть и создал жаберную камеру, в которой располагались kiemenplatten (жаберные тракты). В зависимости от вида, жаберный тракт эвриптерид имел треугольную или овальную форму и, возможно, был приподнят до подушкообразного состояния. На поверхности этого жаберного тракта было несколько шипов (маленьких шипов), что привело к увеличению площади поверхности. Он состоит из губчатой ткани из-за множества инвагинаций в структуре.
Хотя kiemenplatte упоминается как «жаберный тракт», он не обязательно может функционировать как настоящие жабры. У других животных жабры используются для поглощения кислорода из воды и являются отростками стенки тела. Несмотря на то, что эвриптериды явно являются в основном водными животными, которые почти наверняка эволюционировали под водой (некоторые эвриптериды, такие как птериготиды, были бы даже физически неспособны ходить по суше), маловероятно, что жаберный тракт содержал функциональные жабры при сравнении органа с жабрами других беспозвоночные и даже рыбы. Предыдущие интерпретации часто идентифицировали эвриптеридные «жабры» как гомологичные с жабрами других групп (отсюда и терминология), с газообменом, происходящим внутри губчатого тракта, и структурой жаберно-сердечных и дендритных вен (как в родственных группах), несущих насыщенную кислородом кровь в тело. Основной аналогией, использованной в предыдущих исследованиях, были подковообразные крабы, хотя их жаберная структура и структура эвриптерид заметно отличаются. У подковообразных крабов жабры более сложные и состоят из множества пластин (пластин), которые дают большую площадь поверхности, используемую для газообмена. Кроме того, жаберный тракт эвриптерид пропорционально слишком мал, чтобы поддерживать их, если он аналогичен жабрам других групп. Чтобы быть функциональными жабрами, они должны были быть высокоэффективными и потребовать высокоэффективной системы кровообращения. Однако считается маловероятным, что этих факторов было бы достаточно, чтобы объяснить большое несоответствие между размером жаберного тракта и размером тела.
Вместо этого было высказано предположение, что «жаберный тракт» был органом для дыхания воздуха, возможно, фактически являясь легким, пластроном или a. Пластроны - это органы, которые некоторые членистоногие эволюционировали вторично, чтобы дышать воздухом под водой. Это считается маловероятным объяснением, поскольку эвриптериды с самого начала эволюционировали в воде, и у них не было органов, эволюционировавших из имеющихся органов дыхания воздухом. Кроме того, пластроны обычно обнажены на внешних частях тела, а жаберные пути эвриптерид расположены позади блатфюссена. Напротив, среди респираторных органов членистоногих жаберные пути эвриптерид наиболее напоминают псевдотрахеи, обнаруженные у современных изопод. Эти органы, названные псевдотрахеями, из-за некоторого сходства с трахеями (дыхательными путями) дыхательных организмов, похожи на легкие и присутствуют в плеоподах (задних ногах) равноногих. Строение псевдотрахей сравнивали с губчатой структурой жаберных путей эвриптерид. Возможно, эти два органа функционировали одинаково.
Некоторые исследователи предположили, что эвриптериды, возможно, были адаптированы к амфибийному образу жизни, используя всю структуру жаберных путей в качестве жабр и впячиваний внутри них в качестве псевдотрахеи. Однако такой образ жизни мог быть невозможен с физиологической точки зрения, поскольку давление воды заставило бы воду проникнуть в инвагинации, что привело бы к удушью. Более того, большинство эвриптерид всю свою жизнь жили в воде. Независимо от того, сколько времени было проведено на суше, органы дыхания в подводной среде должны были присутствовать. Истинные жабры, которые предположительно располагались внутри жаберной камеры внутри блатфюссена, остаются неизвестными у эвриптерид.
 Личиночный (слева) и ювенильный (справа) возрасты of Strobilopterus (не в масштабе)
Личиночный (слева) и ювенильный (справа) возрасты of Strobilopterus (не в масштабе) Как и все членистоногие, эвриптериды созрели и выросли через статические стадии развития, называемые возрастными стадиями. Эти возрастные стадии перемежались периодами, в течение которых эвриптериды проходили шелушение (линька кутикулы), после чего они подвергались быстрому и немедленному росту. Некоторые членистоногие, например насекомые и многие ракообразные, в процессе созревания претерпевают огромные изменения. Хелицераты, в том числе эвриптериды, обычно считаются прямыми разработчиками, не претерпевающими серьезных изменений после вылупления (хотя дополнительные сегменты тела и дополнительные конечности могут быть получены в течение онтогенеза у некоторых линий, например xiphosurans и морские пауки ). Были ли эвриптериды истинными прямыми разработчиками (птенцы более или менее идентичны взрослым) или гемианаморфными прямыми разработчиками (с дополнительными сегментами и конечностями, которые потенциально могут быть добавлены во время онтогенеза), было спорным в прошлом.
Гемианаморфное прямое развитие имело место. наблюдались у многих групп членистоногих, таких как трилобиты, мегачейры, базальные ракообразные и базальные многоножки. Истинное прямое развитие иногда упоминалось как черта, уникальная для паукообразных. Исследований онтогенеза эвриптерид было мало, так как в летописи окаменелостей практически отсутствуют экземпляры, которые можно было бы с уверенностью сказать, что это молодые особи. Возможно, что многие виды эвриптерид, которые считаются отдельными, на самом деле представляют собой ювенильные экземпляры других видов, при этом палеонтологи редко учитывают влияние онтогенеза при описании новых видов.
Исследования хорошо сохранившегося комплекса ископаемых эвриптерид из Пражская возрастная формация Beartooth Butte в Вайоминг, состоящая из нескольких экземпляров различных стадий развития эвриптерид Jaekelopterus и Strobilopterus, показала, что Онтогенез эвриптерид был более или менее параллелен и похож на онтогенез вымерших и современных ксифосуран, за исключением того, что эвриптериды вылупились с полным набором придатков и опистосомных сегментов. Таким образом, эвриптериды были не гемианаморфными непосредственными разработчиками, а настоящими прямыми разработчиками, такими как современные паукообразные.
Наиболее часто наблюдаемое изменение, происходящее в онтогенезе (за исключением некоторых родов, таких как Eurypterus, которые, по-видимому, были статичными) - это метастома. становится пропорционально менее широким. Это онтогенетическое изменение наблюдалось у представителей нескольких надсемейств, таких как Eurypteroidea, Pterygotioidea и Moselopteroidea.
 Pterygotus охота Биркения.
Pterygotus охота Биркения.Отсутствие ископаемого содержимого кишечника из эвриптериды известны, поэтому прямых доказательств их диеты нет. Биология эвриптерид особенно наводит на мысль о плотоядном образе жизни. Многие не только были крупными (как правило, большинство хищников, как правило, крупнее своей добычи), но и обладали стереоскопическим зрением (способностью воспринимать глубину). Ноги многих эвриптерид были покрыты тонкими шипами, которые использовались как для передвижения, так и для сбора пищи. В некоторых группах эти колючие придатки стали сильно специализированными. У некоторых эвриптерид Carcinosomatoidea обращенные вперед придатки были большими и с чрезвычайно удлиненными шипами (как у Mixopterus и Megalograptus ). У представителей , производных от Pterygotioidea, придатки были полностью без шипов, но вместо них имели специальные когти. Другие эвриптериды, лишенные этих специализированных придатков, вероятно, питались так же, как современные подковообразные крабы, хватали и измельчали пищу своими придатками, прежде чем затолкать ее в рот, используя свои хелицеры.
Окаменелости, сохраняющие пищеварительные тракты. были обнаружены по окаменелостям различных эвриптерид, среди которых Carcinosoma, Acutiramus и Eurypterus. Хотя о потенциальном анальном отверстии сообщалось от тельсона экземпляра Buffalopterus, более вероятно, что задний проход был открыт через тонкую кутикулу между последним сегментом перед тельсоном и сам тельсон, как и у современных подковообразных крабов.
Копролиты эвриптерид, обнаруженные в отложениях ордовикского возраста в Огайо, содержащие фрагменты трилобита и эвриптериды Megalograptus ohioensis в сочетании с полными экземплярами того же вида эвриптерид, предположительно представляют свидетельство каннибализма. Подобные копролиты, относящиеся к виду Lanarkopterus dolichoschelus из ордовика штата Огайо, содержат фрагменты бесчелюстной рыбы и фрагменты более мелких экземпляров самого Lanarkopterus.
Хотя вершина хищника могла бы выполнять роль были ограничены самыми крупными эвриптеридами, более мелкие эвриптериды, вероятно, сами по себе были грозными хищниками, как и их более крупные родственники.
 Тип A Генитальный придаток Adelophthalmus mansfieldi
Тип A Генитальный придаток Adelophthalmus mansfieldi  Тип В генитальный отросток Kokomopterus longicaudatus
Тип В генитальный отросток Kokomopterus longicaudatus Как и во многих других полностью вымерших группах, понимание и исследование воспроизводства и полового диморфизма эвриптерид затруднено, поскольку они известны только по окаменелым раковинам и панцирям. В некоторых случаях может не быть достаточно очевидных различий для разделения полов на основании только морфологии. Иногда два пола одного и того же вида интерпретируются как два разных вида, как это было в случае с двумя видами Drepanopterus (D. bembycoides и D. lobatus).
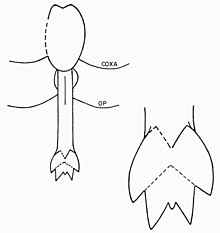
Просома эвриптериды - это состоит из первых шести сегментов экзоскелета, слитых вместе в более крупную структуру. Седьмой сегмент (таким образом, первый опистосомный сегмент) называется метастомой, а восьмой сегмент (отчетливо пластинчатый) называется жаберной крышкой и содержит генитальную апертуру. Нижняя сторона этого сегмента занята генитальной крышечкой - структурой, которая произошла от седьмой и восьмой пары придатков у предков. В его центре, как и у современных подковообразных крабов, находится половой придаток. Этот придаток, удлиненный стержень с внутренним каналом, встречается в двух различных морфах, обычно называемых «типом A» и «типом B». Эти генитальные придатки часто сохраняются в окаменелостях и были предметом различных интерпретаций репродукции эвриптерид и полового диморфизма.
Придатки типа A обычно длиннее, чем у типа B. В некоторых родах они делятся на разные количество секций, например, у Eurypterus, где придаток типа A разделен на три, а придаток типа B - только на два. Такое разделение полового придатка обычное дело у эвриптерид, но число не универсальное; например, придатки обоих типов в семействе Pterygotidae неразделимы. Придаток типа А также вооружен двумя изогнутыми шипами, называемыми фурка (лат. для «вилки»). Также возможно наличие фурки в придатке типа В, и структура может представлять собой несросшиеся кончики придатков. Между дорсальной и вентральной поверхностями блатфуса, связанного с придатками типа А, расположен набор органов, традиционно описываемых как «трубчатые органы» или «роговые органы». Эти органы чаще всего интерпретируются как сперматеки (органы для хранения сперматозоидов ), хотя эта функция еще не доказана окончательно. У членистоногих сперматеки используются для хранения сперматофоров, полученных от самцов. Это означало бы, что придаток типа A - это женский морф, а придаток типа B - мужской. Дополнительным доказательством придатков типа А, представляющих женскую морфу генитальных придатков, является их более сложная конструкция (общая тенденция для гениталий самок членистоногих). Возможно, что большая длина придатка типа A означает, что он использовался в качестве яйцеклада (использовался для откладывания яиц). Различные типы генитальных придатков не обязательно являются единственной особенностью, отличающей половую принадлежность эвриптерид. В зависимости от рассматриваемого рода и вида другие особенности, такие как размер, количество украшений и пропорциональная ширина тела, могут быть результатом полового диморфизма. В целом эвриптериды с придатками типа B (самцы) оказались пропорционально шире, чем эвриптериды с придатками типа A (самки) тех же родов.
Основная функция длинных придатков типа A, предполагаемых самками. скорее всего, будет забирать сперматофор из субстрата в репродуктивный тракт, чем служить яйцекладом, поскольку яйцеклады членистоногих обычно длиннее, чем придатки эвриптерид типа А. Вращая стороны жаберной крышки, можно было бы опустить придаток от тела. Из-за того, что разные пластины накладываются друг на друга в месте его расположения, придаток был бы невозможен без мышечных сокращений, движущихся вокруг жаберной крышки. Он бы оставался на месте, когда им не пользовались. Фурка на придатках типа А могла способствовать разрыву сперматофора, чтобы освободить внутреннюю свободную сперму для поглощения. Считается, что «роговые органы», возможно, сперматеки, были напрямую связаны с придатком через тракты, но эти предполагаемые тракты остаются несохраненными в доступном ископаемом материале.
Придатки типа B, предположительно мужские, могли бы произвести, сохраненный и, возможно, сформированный сперматофор в форме сердца на дорсальной поверхности отростка. Широкое генитальное отверстие позволило бы сразу высвободить большое количество сперматофора. Длинная фурка, связанная с придатками типа B, возможно, способная опускаться, как придаток типа A, могла быть использована для определения того, подходит ли субстрат для отложения сперматофора.
 Реконструкция Pentecopterus, самой ранней известной эвриптериды. Семейство, к которому принадлежит Pentecopterus, Megalograptidae, было первой действительно успешной группой эвриптерид.
Реконструкция Pentecopterus, самой ранней известной эвриптериды. Семейство, к которому принадлежит Pentecopterus, Megalograptidae, было первой действительно успешной группой эвриптерид. До 1882 года эвриптериды не были известны до силурийского периода. Современные открытия, сделанные с 1880-х годов, расширили знания о ранних эвриптеридах ордовикского периода. Самые ранние эвриптериды, известные сегодня, мегалографтид Pentecopterus, относятся к дарривильскому этапу среднего ордовика, 467,3 миллионов лет назад. Имеются также сообщения о еще более ранних ископаемых эвриптеридах в отложениях позднего тремадока (раннего ордовика) в Марокко, но они еще не были полностью изучены.
Pentecopterus был относительно производным эвриптеридом, частью семейства мегалографтидов в суперсемействе карциносоматоидов. Его производное положение предполагает, что большинство клад эвриптерид, по крайней мере в пределах подотряда эвриптерид, уже были установлены в этот момент в течение среднего ордовика. Самая ранняя известная стилонуриновая эвриптерида, Brachyopterus, также имеет возраст среднего ордовика. Присутствие представителей обоих подотрядов указывает на то, что примитивные стеблевые эвриптериды предшествовали им, хотя они пока неизвестны в летописи окаменелостей. Присутствие нескольких клад эвриптерид в среднем ордовике предполагает, что либо эвриптериды возникли в раннем ордовике и испытали быстрое и взрывное излучение и диверсификацию вскоре после появления первых форм, либо что группа возникла намного раньше, возможно, во время кембрия . период.
Таким образом, точное время происхождения эвриптериды остается неизвестным. Хотя окаменелости, называемые «примитивными эвриптеридами», иногда описывались из отложений кембрийского или даже докембрийского возраста, сегодня они не признаются как эвриптериды, а иногда даже не как родственные формы. Некоторые животные, ранее считавшиеся примитивными эвриптеридами, такие как род Strabops из кембрия Миссури, теперь классифицируются как агласпидиды или страбопиды. Агласпидиды, которые когда-то считались примитивными хелицератами, теперь рассматриваются как группа, более тесно связанная с трилобитами.
Летопись окаменелостей ордовикских эвриптерид довольно скудна. Большинство эвриптерид, когда-то известных из ордовика, с тех пор оказались ложными отождествлениями или псевдокаменелостями. Сегодня только 11 видов можно с уверенностью идентифицировать как представителей ордовикских эвриптерид. Эти таксоны делятся на две отдельные экологические категории; крупные и активные хищники с древнего континента Лаурентия и демерсальные (живущие на морском дне ) и базальные животные с континентов Авалония и Гондвана. Лаврентийские хищники, отнесенные к семейству Megalograptidae (под угрозой роды Echinognathus, Megalograptus и Pentecopterus), вероятно, представляют первую действительно успешную группу эвриптерид, испытавшую небольшую радиацию в позднем ордовике. 199>Силурийский Реконструкция Erettopterus, представителя очень успешных силурийских и девонских семейств эвриптерид Pterygotidae
Реконструкция Erettopterus, представителя очень успешных силурийских и девонских семейств эвриптерид Pterygotidae
Эвриптериды были самыми разнообразными и многочисленны между средним силуром и ранним девоном, с абсолютным пиком в разнообразие во время эпохи Придоли, 423–419,2 миллиона лет назад, самого позднего силурия. Этот пик разнообразия был признан с начала двадцатого века; из примерно 150 видов эвриптерид, известных в 1916 году, более половины происходили из силурия, а треть - только из позднего силура.
Хотя стилонуриновые эвриптериды в целом оставались редкими и малочисленными, как и раньше. Во время предшествующего ордовика численность и разнообразие эвриптериновых эвриптерид резко возросли. В большинстве ископаемых пластов силурия эвриптеридные эвриптериды составляют 90% всех присутствующих эвриптерид. Хотя некоторые из них, вероятно, уже присутствовали в позднем ордовике (до сих пор просто отсутствовали в летописи окаменелостей), подавляющее большинство групп эвриптерид впервые зарегистрированы в слоях силурийского возраста. К ним относятся как группы стилонурина, такие как Stylonuroidea, Kokomopteroidea и Mycteropoidea, так и группы эвриптерина, такие как Pterygotioidea, Eurypteroidea и Waeringopteroidea.
Самым успешным эвриптеридом был средне- и позднесилурийский Eurypterus, универсал, с равной вероятностью участвовавший в хищничестве или падальщике. Считалось, что охотились в основном на мелких беспозвоночных с мягким телом, таких как черви, виды этого рода (из которых наиболее распространен типовой вид E. remipes) составляют более 90% (возможно, поскольку около 95%) всех известных ископаемых экземпляров эвриптерид. Несмотря на их огромное количество, Eurypterus известны только из относительно короткого временного диапазона, впервые появившись в конце эпохи Лландовери (около 432 миллионов лет назад) и вымершие к концу эпохи Придоли. Eurypterus также был ограничен второстепенным суперконтинентом Euramerica (состоящим из экваториальных континентов Avalonia, Baltica и Laurentia), который был полностью колонизирован этим видом во время его слияния и не смог пересечь обширные просторы океана, отделяющие этот континент от других частей мира, таких как южный суперконтинент Гондвана. Таким образом, Eurypterus был географически ограничен береговой линией и мелководными внутренними морями Еврамерики.
В течение позднего силура птеригидные эвриптериды, большие и специализированные формы с несколькими новыми приспособлениями, такими как большие и плоские тельсоны, которые можно было использовать в качестве рулей появились большие и специализированные хелицеры с увеличенными клешнями для управления (и, возможно, в некоторых случаях для убийства) добычи. Хотя самые крупные представители семейства появились в девоне, большие двухметровые птеригиды, такие как Acutiramus, уже существовали в позднем силурии. Их экология варьировалась от общего хищнического поведения до хищничества из засад, а некоторые, такие как сам Pterygotus, были активными высшими хищниками в морских экосистемах позднего силура. Птериготиды также, очевидно, были способны пересекать океаны, став одной из двух групп эвриптерид, достигших космополитического распространения.
 Реконструкция аделофтальма, единственного эвриптерина (с плавательными веслами) эвриптериды, чтобы пережить позднедевонское вымирание и сохраниться в последующие каменноугольный и пермский периоды.
Реконструкция аделофтальма, единственного эвриптерина (с плавательными веслами) эвриптериды, чтобы пережить позднедевонское вымирание и сохраниться в последующие каменноугольный и пермский периоды. Хотя эвриптериды продолжали существовать. многочисленная и разнообразная в течение раннего девона (например, приведшая к развитию pterygotid Jaekelopterus, самого крупного из всех членистоногих), эта группа была одной из многих, сильно пострадавших от позднедевонского вымирания. Событие вымирания, которое, как известно, затронуло только морскую жизнь (особенно трилобиты, брахиоподы и рифовые организмы), эффективно нанесло ущерб численности и разнообразию, ранее наблюдавшимся у эвриптерид.
Существенное сокращение разнообразия уже началось в раннем девоне, и эвриптериды были редкостью в морской среде к позднему девону. Во время франской стадии вымерли четыре семьи, а на более поздней фаменской стадии вымерли еще пять семейств. Поскольку больше всего пострадали морские группы, эвриптериды в первую очередь пострадали в рамках подотряда эвриптеридов. Только одна группа stylonurines (семейство Parastylonuridae ) вымерла в раннем девоне. Всего лишь два семейства эвриптерин дожили до позднего девона (Adelophthalmidae и Waeringopteridae). Наиболее значительный упадок эвриптерин произошел в раннем девоне, во время которого более 50% их разнообразия было потеряно всего за 10 миллионов лет. С другой стороны, стилонурины сохранялись на протяжении всего периода с более или менее постоянным разнообразием и численностью, но были затронуты в течение позднего девона, когда многие из более старых групп были заменены новыми формами в семействах Mycteroptidae и Hibbertopteridae.
Возможно, что модели катастрофического вымирания, наблюдаемые в подотряде эвриптерин, были связаны с появлением более производных рыб. Упадок эвриптеринов начался в тот момент, когда бесчелюстные рыбы впервые стали более развитыми, и совпадает с появлением плакодерм (панцирных рыб) как в Северной Америке, так и в Европе.
Стилонурины выживших гиббертоптеридов и миктероптидов. семьи полностью избегали конкуренции с рыбой, развиваясь в направлении новой и отчетливой экологической ниши. Эти семьи испытали радиацию и диверсификацию в течение позднего девона и раннего карбона, последнюю радиацию в пределах эвриптерид, которая дала начало нескольким новым формам, способным к «подметанию» (рывку субстрата в поисках добычи).
 Реконструкция иббертоптерида Campylocephalus, последней известной выжившей эвриптериды. Campylocephalus вымерли во время пермско-триасового вымирания или незадолго до этого.
Реконструкция иббертоптерида Campylocephalus, последней известной выжившей эвриптериды. Campylocephalus вымерли во время пермско-триасового вымирания или незадолго до этого. Только три семейства эвриптерид - Adelophthalmidae, Hibbertopteridae и Mycteroptidae - полностью пережили вымирание. Все это были пресноводные животные, из-за чего эвриптериды вымерли в морской среде. После исчезновения морских хищников эвриптерид саркоптерийские рыбы, такие как ризодонты, стали новыми высшими хищниками в морской среде. Единственное сохранившееся семейство эвриптеринов, Adelophthalmidae, было представлено только одним родом Adelophthalmus. Гиббертоптериды, миктероптиды и Adelophthalmus дожили до пермского периода.
Adelophthalmus стал наиболее распространенным из всех позднепалеозойских эвриптеридов, существовавших в большем количестве и разнообразии, чем выжившие стилонурины, и разносторонне развивавшихся в отсутствие других эвриптеринов. Из 33 видов, относящихся к Adelophthalmus, 23 (69%) происходят только из карбона. Пика своего разнообразия род достиг в позднем карбоне. Хотя аделофтальм уже был относительно широко распространен и представлен на всех основных массивах суши в позднем девоне, объединение Пангеи в глобальный суперконтинент в течение последних двух периодов палеозоя позволило Аделофтальму получить почти всемирное признание.
В течение позднего карбона и ранней перми Adelophthalmus был широко распространен, обитая в основном в солоноватой и пресноводной среде, прилегающей к прибрежным равнинам. Эти среды поддерживались благоприятными климатическими условиями. Они не сохранились, поскольку изменения климата из-за образования Пангеи изменили структуру осадконакопления и растительность по всему миру. С исчезновением среды обитания Adelophthalmus сократилось в количестве и уже вымерло к леонардианской стадии ранней перми.
Mycteroptids и hibbertopterids продолжали выживать в течение некоторого времени, по одному роду каждого группа, известная из пермских слоев: Hastimima и Campylocephalus соответственно. Хастимима вымерла в ранней перми, как и Adelophthalmus, в то время как Campylocephalus существовал дольше. Массивный неполный панцирь из отложений поздней перми (чансин ) в России представляет собой единственные ископаемые останки вида C. permianus, которые могли достигать 1,4 метра (4,6 фута) в длину. Этот гигант был последней выжившей эвриптеридой. Эвриптериды из ископаемых пластов выше перми не известны. Это указывает на то, что последние эвриптериды погибли либо в результате катастрофического вымирания в его конце, либо в какой-то момент незадолго до него. Это событие вымирания, пермско-триасовое вымирание, является самым разрушительным массовым вымиранием из зарегистрированных, и привело к исчезновению многих других успешных палеозойских групп, таких как трилобиты.
 Рисунок Eurypterus напоминает Джеймса Э. Де Кея (1825).
Рисунок Eurypterus напоминает Джеймса Э. Де Кея (1825). Первый известный образец эвриптерид был обнаружен в породах силурийского возраста из Нью-Йорка, и по сей день это одно из самых богатых мест окаменелостей эвриптерид. Сэмюэл Л. Митчилл описал образец, обнаруженный около Вестморленда в графстве Онейда в 1818 году. Он ошибочно определил окаменелость как образец рыбы Silurus, вероятно, из-за странного, сома внешнего вида панциря. Семь лет спустя, в 1825 году, Джеймс Э. ДеКей исследовал окаменелость и определил, что она явно принадлежит членистоногому. Он думал, что окаменелость, которую он назвал Eurypterus remipes, представляет ракообразных отряда Branchiopoda, и предположил, что это могло быть недостающее звено между трилобитами и более производными жабероногими моллюсками. Имя Eurypterus происходит от древнегреческих слов εὐρύς (eurús), означающих «широкий» или «широкий», и πτερόν (pteron), означающего «крыло».
В 1843 г., Герман Бурмейстер опубликовал свой взгляд на таксономию трилобитов и то, как эта группа связана с другими организмами, живыми и вымершими, в работе Die Organization der Trilobiten aus ihren lebenden Verwandten entwickelt. Он считал трилобитов ракообразными, как и предыдущие авторы, и классифицировал их вместе с тем, что он считал их ближайшими родственниками, Eurypterus и родом, в кладу, которую он назвал «Palaeadae». В Палеадах Бурмейстер построил три семьи; «Trilobitae» (состоящий из всех трилобитов), «Cytherinidae» (состоящий только из Cytherina, животное, которое сегодня рассматривается как остракод ) и Eurypteridae (состоящий из Eurypterus, затем включая три вида).
Четвертый род эвриптерид, подлежащий описанию (после Hibbertopterus в 1836 г. и Campylocephalus в 1838 г., не идентифицированных как эвриптериды до позднего времени), из тех, которые все еще рассматриваются как таксономически обоснованные в наше время это был Птеригот, описанный Луи Агассисом в 1839 году. Птеригот был значительно больше по размеру, чем Евриптер, и когда первые ископаемые были обнаружены каменоломнями в Шотландии, они назывались «Серафимы » рабочими. Сначала Агассис подумал, что окаменелости представляют собой останки рыб, а имя Pterygotus означает «крылатая рыба», и осознал их природу как останки членистоногих только через пять лет, в 1844 году.
В 1849 году Фредерик Маккой классифицировал Pterygotus вместе с Eurypterus и Bellinurus (род, сегодня рассматриваемый как ксифосуран) в составе Eurypteridae Бурмейстера. Маккой считал Eurypteridae группой ракообразных в пределах отряда Entomostraca, тесно связанных с подковообразными крабами. Четвертый род, Slimonia, основанный на ископаемых останках, ранее отнесенных к новому виду Pterygotus, был отнесен к Eurypteridae в 1856 году Дэвидом Пейджем.
 Древо эволюции эвриптерид, как и предполагалось. Авторы Джон Мейсон Кларк и Рудольф Рюдеманн в 1912 году.
Древо эволюции эвриптерид, как и предполагалось. Авторы Джон Мейсон Кларк и Рудольф Рюдеманн в 1912 году. Книга Яна Нешковского De Euryptero Remipede (1858) содержала подробное описание Eurypterus fischeri (теперь рассматриваемого как синоним другого вида Eurypterus, E. tetragonophthalmus), который, наряду с монографией «О роду Pterygotus», написанной Томасом Генри Хаксли и Джоном Уильямом Солтером, и исчерпывающим описанием различных эвриптерид Нью-Йорка в томе 3 «Палеонтологии Нью-Йорка» (1859 г.) Джеймса Холла внес большой вклад в понимание разнообразия и биологии эвриптерид. Эти публикации были первыми, кто полностью описал всю анатомию эвриптерид, признав полное количество просомальных придатков и количество преабдоминальных и постабдоминальных сегментов. И Нешковский, и Холл признали, что эвриптериды были тесно связаны с современными хелицератами, такими как подковообразные крабы.
В 1865 году Генри Вудворд описал род Stylonurus (названный и фигурировал, но не полностью описал Дэвидом Пейджем в 1856 г.) и поднял ранг Eurypteridae до порядкового, фактически создав Eurypterida в качестве таксономической единицы, которой она считается сегодня. В работе «Анатомия и отношения эвриптерид» (1893) Малкольм Лори значительно расширил знания и обсуждение анатомии эвриптерид и их взаимосвязей. Он сосредоточился на том, как эвриптериды связаны друг с другом и с трилобитами, ракообразными, скорпионами, другими паукообразными и подковообразными крабами. Описание Eurypterus fischeri Герхардом Хольмом в 1896 году было настолько подробным, что этот вид стал одним из наиболее известных среди всех вымерших животных, настолько, что знания E. fischeri были сопоставимы со знаниями его современных родственников (таких как атлантический подковообразный краб ). Описание также помогло укрепить тесную связь между эвриптеридами и другими хелицератами, продемонстрировав многочисленные гомологии между двумя группами.
В 1912 году Джон Мейсон Кларк и Рудольф Рюдеманн опубликовал Eurypterida of New York, в которой обсуждались все виды эвриптерид, извлеченные из ископаемых отложений. Кларк и Рюдеманн создали одно из первых филогенетических деревьев эвриптерид, разделив отряд на два семейства; Eurypteridae (отличаются гладкими глазами и включают Eurypterus, Anthraconectes, Stylonurus, Eusarcus, Dolichopterus, Onychopterus и Drepanopterus) и Pterygotidae (выделенные по фасеточным глазам, включая Pterygotus, Erettopterus, Slimonia и Hughmilleria ). Считалось, что обе семьи произошли от общего предка Страбопов. Как и предыдущие авторы, Кларк и Рудеманн также поддерживали тесную связь между эвриптеридами и подковообразными крабами (объединенными в класс Merostomata), но также обсуждали альтернативные гипотезы, такие как более близкое родство с паукообразными
.
 Эвриптериды исторически считались тесно связанными с ксифосуранами, такими как атлантический подковообразный краб (вверху), объединенным в класс Merostomata. Недавние исследования свидетельствуют в пользу более тесного родства с паукообразными, такими как Heterophrynus (внизу), объединенными в кладу Sclerophorata.
Эвриптериды исторически считались тесно связанными с ксифосуранами, такими как атлантический подковообразный краб (вверху), объединенным в класс Merostomata. Недавние исследования свидетельствуют в пользу более тесного родства с паукообразными, такими как Heterophrynus (внизу), объединенными в кладу Sclerophorata.Исторически сложилась тесная связь между эвриптеридами и ксифосуранами. (таких как современный атлантический подковообразный краб) предполагалось большинством исследователей. Некоторые гомологии поддерживают эту точку зрения, например, коррелирующие сегменты придатков и просомы. Кроме того, наличие пластинчатых придатков, несущих «жаберные тракты» на придатках опистосомы (блатфюссена), было упомянуто как важная гомология. В последние несколько десятилетий девятнадцатого века были установлены дальнейшие гомологии, такие как схожие структуры сложных глаз птеригота и подковообразных крабов (считавшиеся особенно решающими, поскольку глаз подковообразного краба считался обладающим почти уникальной структурой) и сходство в онтогенезе внутри обеих групп. Это онтогенетическое сходство было замечено как наиболее очевидное при изучении непионических стадий (стадия развития сразу после эмбриональной стадии) в обеих группах, во время которых и ксифосуран, и эвриптериды имеют пропорционально больший панцирь, чем взрослые особи, обычно более широкие., обладают отчетливым гребнем посередине, имеют меньшее количество сегментов, которые не дифференцируются и имеют недоразвитый тельсон.
Из-за этих сходств ксифосураны и эвриптериды часто объединяются в один класс или подкласс, названный Merostomata (возведен для размещения обеих групп Генри Вудвордом в 1866 году). Хотя ксифосураны (как и эвриптериды) исторически считались ракообразными из-за их дыхательной системы и водного образа жизни, эта гипотеза была дискредитирована после того, как были обнаружены многочисленные сходства между подковообразными крабами и паукообразными. Некоторые авторы, такие как Джон Стерлинг Кингсли в 1894 году, классифицировали Merostomata как сестринскую группу по отношению к Arachnida в класс «Acerata» в подтипе «Branchiata». Другие, такие как Рэй Ланкестер в 1909 году, пошли дальше и классифицировали Merostomata как подкласс внутри Arachnida, возведенный в ранг класса.
В 1866 году Эрнст Геккель классифицировал Merostomata (содержащие фактически только Eurypterida) и Xiphosura в группу, которую он назвал Gigantostraca среди ракообразных. Хотя Геккель не определил какой-либо таксономический ранг для этой клады, более поздние исследователи, такие как Джон Стерлинг Кинзсли, интерпретировали его как эквивалент ранга подкласса, такого как Malacostraca и Entomostraca. В последующих исследованиях Gigantostraca рассматривалась как синоним Merostomata (редко) и самой Eurypterida (чаще).
 Восстановление примитивного кембрийского арахноморфа Fuxianhuia, один из самых базальных членов линии, который в конечном итоге приведет к хелицератам и трилобитам.
Восстановление примитивного кембрийского арахноморфа Fuxianhuia, один из самых базальных членов линии, который в конечном итоге приведет к хелицератам и трилобитам.Филогенетический анализ (результаты представлены на кладограмме ниже), проведенного Джеймсом Ламсделлом в 2013 году, о взаимоотношениях внутри Xiphosura и отношениях с другими близкородственными группами (включая эвриптериды, которые были представлены в анализе родами Eurypterus, Parastylonurus, Rhenopterus и Stoermeropterus ) пришел к выводу, что ксифосура в современном понимании была парафилетической (группа, имеющая последнего общего предка, но не включающая всех потомков этого предка) и, таким образом, не действительна филогенетическая группа. Эвриптериды были обнаружены как близкородственные паукообразным, а не ксифосуранам, образуя группу Sclerophorata внутри клады Dekatriata (состоящую из склерофоратов и chasmataspidids ). Ламсделл отметил, что возможно, что Dekatriata является синонимом Sclerophorata, поскольку репродуктивная система, основная определяющая характеристика склерофоратов, не была полностью изучена у chasmataspidids. Dekatriata, в свою очередь, является частью Prosomapoda, группы, включающей Xiphosurida (единственная монофилетическая группа ксифосуранов) и другие родовые роды.
| Arachnomorpha |
|

