
Войти
| Вирус гаммагерпеса человека 4 | |
|---|---|
 | |
| Электронная микрофотография двух вирионов Эпштейна – Барра (вирусные частицы) с круглыми капсидами, свободно окруженными мембраной оболочкой | |
| Классификация вирусов | |
| (без ранжирования): | Вирус |
| Область: | Дуплоднавирия |
| Царство: | Heunggongvirae |
| Тип: | Peploviricota |
| Класс: | Herviviricetes |
| Порядок: | Herpesvirales |
| Семья: | Herpesviridae |
| Род: | Лимфокриптовирус |
| Виды: | Человеческий гаммагерпесвирус 4 |
| Синонимы | |
| |
вирус Эпштейна – Барра (EBV ), формально называемый гаммагерпесвирус человека 4, является одним из девяти известные типы вирусов герпеса человека в семействе герпеса и являются одним из наиболее распространенных вирусов у людей.
Он наиболее известен как причина инфекционного мононуклеоза («моно» или «железистая лихорадка»). Он также связан с различными доброкачественными, предраковыми и злокачественными лимфопролиферативными заболеваниями, связанными с вирусом Эпштейна-Барра, такими как лимфома Беркитта, гемофагоцитарный лимфогистиоцитоз и лимфома Ходжкина ; нелимфоидные злокачественные новообразования, такие как рак желудка и карцинома носоглотки ; и состояния, связанные с вирусом иммунодефицита человека, такие как волосатая лейкоплакия и лимфомы центральной нервной системы лимфомы. Вирус также связан с детскими расстройствами синдромом Алисы в стране чудес и острой мозжечковой атаксией и, по некоторым данным, с более высоким риском развития некоторых аутоиммунных заболеваний, особенно дерматомиозит, системная красная волчанка, ревматоидный артрит, синдром Шегрена и рассеянный склероз. Считается, что около 200 000 случаев рака в год связаны с EBV.
Заражение EBV происходит при оральном переносе слюны и генитальных выделений.
Большинство людей заражаются EBV и приобретают адаптивный иммунитет. В Соединенных Штатах около половины всех пятилетних детей и около 90% взрослых имеют признаки перенесенной инфекции. Младенцы становятся восприимчивыми к ВЭБ, как только исчезает защита материнскими антителами. Многие дети заражаются ВЭБ, и эти инфекции обычно не вызывают симптомов или неотличимы от других легких и кратковременных детских болезней. В Соединенных Штатах и других развитых странах многие люди не инфицированы ВЭБ в детстве. Когда инфицирование EBV происходит в подростковом возрасте, это вызывает инфекционный мононуклеоз в 35–50% случаев.
EBV заражает B-клетки иммунной системы и эпителиальные клетки. После того как первоначальная литическая инфекция EBV взята под контроль, EBV латентность сохраняется в B-клетках человека на всю оставшуюся жизнь.
У детей, инфицированных ВЭБ, мало симптомов или они могут даже проявляться бессимптомно, но когда ВЭБ заразился подростком или взрослым, это может вызвать усталость, лихорадку, воспаление горла, опухоль лимфатических узлов в шее, увеличенная селезенка, опухшая печень или сыпь.. Постинфекционные хроническая усталость S Индром также был связан с инфекцией Эпштейна-Барра.
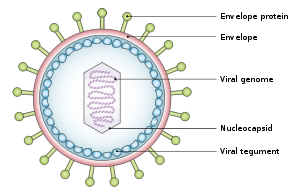 Упрощенная диаграмма структуры EBV
Упрощенная диаграмма структуры EBV Вирус имеет диаметр около 122–180 нм и состоит из двойной спирали дезоксирибонуклеиновой кислоты (ДНК), которая содержит около 172000 пар оснований и 85 гены. ДНК окружена белком нуклеокапсидом, который окружен тегументом, состоящим из белка, который, в свою очередь, окружен оболочкой, содержащей оба липидов и поверхностных выступов гликопротеинов, которые необходимы для инфекции клетки-хозяина. В июле 2020 года группа исследователей сообщила о первой полной атомной модели нуклеокапсида вируса. Эта «первая полная атомная модель [включает] икосаэдрический капсид, связанный с капсидом тегументный комплекс (CATC) и додекамерный портал - аппарат транслокации вирусного генома».
Термин вирусный тропизм относится к типам клеток, которыми заражает EBV. EBV может инфицировать различные типы клеток, включая B-клетки и эпителиальные клетки.
Вирусные трехкомпонентные гликопротеиновые комплексы опосредуют слияние мембран B-клеток; хотя двухкомпонентные комплексы gHgL опосредуют слияние мембран эпителиальных клеток. EBV, которые образуются в B-клетках, имеют низкое количество комплексов gHgLgp42, потому что эти трехкомпонентные комплексы взаимодействуют с молекулами человеческий лейкоцит-антиген класс II, присутствующими в B-клетках в эндоплазматическом reticulum и деградированы. Напротив, EBV из эпителиальных клеток богаты трехкомпонентными комплексами, поскольку эти клетки обычно не содержат молекул HLA класса II. Как следствие, ВЭБ, полученный из В-клеток, более заразен для эпителиальных клеток, а ВЭБ, полученный из эпителиальных клеток, более заразен для В-клеток. Вирусы, у которых отсутствует эта часть, способны связываться с В-клетками человека, но не могут инфицировать.
 Цикл репликации EBV
Цикл репликации EBV EBV может инфицировать оба В-клетки и эпителиальные клетки. Механизмы проникновения в эти две клетки различны.
Чтобы проникнуть в В-клетки, вирусный гликопротеин связывается с клеточным рецептором CD21 (также известным как CR2). Затем вирусный гликопротеин gp42 взаимодействует с клеточными молекулами MHC класса II. Это запускает слияние вирусной оболочки с клеточной мембраной, позволяя EBV проникать в B-клетку. CD35 человека, также известный как рецептор комплемента 1 (CR1), является дополнительным фактором прикрепления для gp350 / 220 и может обеспечивать путь проникновения EBV в CD21-негативные клетки, включая незрелые B-клетки. Инфекция EBV подавляет экспрессию CD35.
Чтобы проникнуть в эпителиальные клетки, вирусный белок BMRF-2 взаимодействует с клеточными β1 интегринами. Затем вирусный белок gH / gL взаимодействует с клеточными / интегринами. Это запускает слияние вирусной оболочки с мембраной эпителиальной клетки, позволяя EBV проникать в эпителиальную клетку. В отличие от входа в B-клетки, входу в эпителиальные клетки фактически препятствует вирусный гликопротеин gp42.
Как только EBV попадает в клетку, вирусный капсид растворяется, и вирусный геном переносится в ядро клетки.
литический цикл, или продуктивная инфекция, приводит к образованию инфекционных вирионов. EBV может подвергаться литической репликации как в B-клетках, так и в эпителиальных клетках. В В-клетках литическая репликация обычно происходит только после реактивации с латентности. В эпителиальных клетках литическая репликация часто непосредственно следует за проникновением вируса.
Чтобы происходила литическая репликация, вирусный геном должен быть линейным. Скрытый геном EBV является кольцевым, поэтому он должен линеаризоваться в процессе литической реактивации. Во время литической репликации вирусная ДНК-полимераза отвечает за копирование вирусного генома. Это контрастирует с латентным периодом, когда ДНК-полимераза клетки-хозяина копирует вирусный геном.
Продукты литического гена продуцируются на трех последовательных стадиях: непосредственно-ранний, ранний и поздний. Продукты литических генов немедленного раннего действия действуют как трансактиваторы, усиливая экспрессию более поздних литических генов. Продукты литического гена для немедленного начала включают BZLF1 (также известный как Zta, EB1, связанный с его геном-продуктом ZEBRA) и (связанный с его геном-продуктом Rta). Продукты ранних литических генов имеют гораздо больше функций, таких как репликация, метаболизм и блокада процессинга антигена. Продукты ранних литических генов включают. Наконец, продукты поздних литических генов, как правило, представляют собой белки со структурными функциями, такие как VCA, который формирует вирусный капсид. Другие продукты позднего литического гена, такие как BCRF1, помогают ВЭБ ускользать от иммунной системы.
EGCG, полифенол в зеленом чае, в исследовании показал, что он подавляет спонтанную литическую инфекцию ВЭБ в ДНК, транскрипция генов и уровни белка в зависимости от времени и дозы ; экспрессия литических генов EBV Zta, Rta и раннего антигенного комплекса EA-D (индуцируемого Rta), однако высокостабильный ген EBNA-1, обнаруживаемый на всех стадиях инфицирования EBV, не изменяется. Специфические ингибиторы (пути) предполагают, что путь Ras / MEK / MAPK способствует литической инфекции EBV через BZLF1 и PI3-K путь через BRLF1, последний полностью отменяет способность BRLF1 аденовирусный вектор для индукции литической формы EBV-инфекции. Кроме того, активация некоторых генов, но не других, изучается, чтобы определить, как вызвать иммунную деструкцию латентно инфицированных В-клеток с помощью TPA или бутирата натрия.
В отличие от литической репликации, латентность не приводит к образованию вирионов. Вместо этого кольцевая ДНК генома EBV находится в клеточном ядре как эписома и копируется клеточной ДНК-полимеразой. В латентном состоянии только часть генов EBV экспрессируется. Скрытый ВЭБ экспрессирует свои гены в одном из трех паттернов, известных как латентные программы. ВЭБ может латентно сохраняться в В-клетках и эпителиальных клетках, но в этих двух типах клеток возможны разные программы латентности.
EBV может отображать одну из трех программ задержки: Latency I, Latency II или Latency III. Каждая программа латентности приводит к продукции ограниченного, отличного набора вирусных белков и вирусных РНК.
| Экспрессия гена | EBNA-1 | EBNA-2 | EBNA- 3 A | EBNA-3 B | EBNA-3 C | EBNA-LP | LMP-1 | LMP-2 A | LMP-2 B | EBER |
|---|---|---|---|---|---|---|---|---|---|---|
| Продукт | Белок | Белок | Белок | Белок | Белок | Белок | Белок | Белок | Белок | нкРНК |
| Латентность I | + | – | – | – | – | – | – | – | – | + |
| Латентность II | + | – | – | – | – | + | + | + | + | + |
| Латентность III | + | + | + | + | + | + | + | + | + | + |
Кроме того, постулируется программа, в которой полностью отключена экспрессия вирусных белков (Задержка 0).
Внутри B-клеток возможны все три программы задержки. Латентность EBV в B-клетках обычно прогрессирует от латентности III к латентности II и к латентности I. Каждый этап латентности однозначно влияет на поведение B-клеток. После заражения покоящихся наивных B-клеток EBV переходит в латентный период III. Набор белков и РНК, продуцируемых в Latency III, трансформирует B-клетку в пролиферирующий бласт (также известный как активация B-клеток). Позже вирус ограничивает экспрессию своего гена и переходит в латентность II. Более ограниченный набор белков и РНК, продуцируемых в Latency II, побуждает B-клетку дифференцироваться в B-клетку памяти. Наконец, EBV еще больше ограничивает экспрессию гена и входит в латентность I. Экспрессия EBNA-1 позволяет геному EBV реплицироваться при делении В-клетки памяти.
Внутри эпителиальных клеток возможна только латентность II.
При первичной инфекции EBV реплицируется в эпителиальных клетках ротоглотки и вызывает латентность III, II и I инфекции в B-лимфоцитах. Скрытая EBV-инфекция В-лимфоцитов необходима для персистенции вируса, последующей репликации в эпителиальных клетках и выделения инфекционного вируса в слюну. Инфекция В-лимфоцитов с латентностью III и II EBV, инфекция с латентностью II эпителиальных клеток полости рта и инфекция NK- или Т-клеток с латентностью II может привести к злокачественным новообразованиям, отмеченным однородным присутствием генома EBV и экспрессией генов.
Скрытый EBV в B-клетках можно реактивировать, чтобы переключиться на литическую репликацию. Известно, что это происходит in vivo, но точно не известно, что это вызывает. In vitro латентный EBV в B-клетках может быть реактивирован путем стимуляции рецептора B-клеток, поэтому реактивация in vivo, вероятно, происходит, когда латентно инфицированные B-клетки реагируют на несвязанные инфекции. In vitro, латентный EBV в B-клетках также можно реактивировать путем обработки клеток бутиратом натрия или 12-O-тетрадеканоилфорбол-13-ацетатом.
Когда ВЭБ инфицирует В-клетки in vitro, в конечном итоге появляются лимфобластоидные клеточные линии, способные к неограниченному росту. Трансформация роста этих клеточных линий является следствием экспрессии вирусного белка.
EBNA-2, EBNA-3C и LMP-1 необходимы для трансформации, тогда как EBNA-LP и EBER - нет.
Считается, что после естественного заражения EBV вирус выполнять некоторые или все из своего репертуара программ экспрессии генов для установления стойкой инфекции. Учитывая исходное отсутствие иммунитета хозяина, литический цикл производит большое количество вирионов для инфицирования других (предположительно) В-лимфоцитов внутри хозяина.
Скрытые программы перепрограммируют и разрушают инфицированные B-лимфоциты для пролиферации и доставки инфицированных клеток к участкам, в которых предположительно сохраняется вирус. В конце концов, когда иммунитет хозяина развивается, вирус сохраняется, отключая большую часть (или, возможно, все) своих генов, лишь изредка реактивируясь с образованием свежих вирионов. В конечном итоге достигается баланс между периодической реактивацией вируса и иммунным надзором хозяина, удаляющим клетки, которые активируют экспрессию вирусных генов.
Местом сохранения EBV может быть костный мозг. EBV-положительные пациенты, у которых собственный костный мозг заменен костным мозгом от EBV-отрицательного донора, оказываются EBV-отрицательными после трансплантации.
Вырабатываются все ядерные белки EBV путем альтернативного сплайсинга транскрипта, начиная с промоторов Cp или Wp на левом конце генома (в общепринятой номенклатуре). Гены упорядочены в геноме EBNA-LP / EBNA-2 / EBNA-3A / EBNA-3B / EBNA-3C / EBNA-1.
Инициирование кодона кодирующей области EBNA-LP создается альтернативным сплайсингом транскрипта ядерного белка. В отсутствие этого инициирующего кодона, EBNA-2 / EBNA-3A / EBNA-3B / EBNA-3C / EBNA-1 будет экспрессироваться в зависимости от того, какой из этих генов альтернативно сплайсирован в транскрипт.
| Белок / ген / антиген | Стадия | Описание |
|---|---|---|
| EBNA-1 | латентный + литический | EBNA -1 белок связывается с ориджином репликации (oriP) в вирусном геноме и опосредует репликацию и разделение эписомы во время деления клетки-хозяина. Это единственный вирусный белок, экспрессируемый в латентный период группы I. |
| EBNA-2 | латентный + литический | EBNA-2 - основной вирусный трансактиватор. |
| EBNA-3 | латентный + литический | Эти гены также связываются с белком RBP-Jκ хозяина. |
| LMP-1 | латентный | LMP-1 представляет собой шестичленный трансмембранный белок, который также важен для трансформации роста, опосредованной EBV. |
| LMP-2 | латентный | LMP-2A / LMP-2B представляют собой трансмембранные белки, которые действуют, блокируя передачу сигналов тирозинкиназы. |
| EBER | латентный | EBER-1 / EBER-2 - это небольшие ядерные РНК, которые связываются с определенными частицами нуклеопротеидов, обеспечивая связывание с PKR (дцРНК-зависимой серин / треониновой протеинкиназой), тем самым ингибируя его функция. EBER являются наиболее распространенными продуктами EBV, транскрибируемыми в инфицированных EBV клетках. Они обычно используются в качестве мишеней для обнаружения EBV в гистологических тканях. ER-частицы также вызывают выработку IL-10, который усиливает рост и ингибирует цитотоксические Т-клетки. |
| v-snoRNA1 | латентный | snoRNA1 вируса Эпштейна-Барра представляет собой коробчатую CD-snoRNA, генерируемую вирусом во время латентного периода. V-snoRNA1 может действовать как miRNA-подобный предшественник, который процессируется в фрагменты РНК размером 24 нуклеотида, которые нацелены на 3'UTR мРНК вирусной ДНК-полимеразы. |
| ebv-sisRNA | латентный | Ebv- sisRNA-1 представляет собой стабильную интронную последовательность РНК, генерируемую во время латентной программы III. После EBER, это третья по численности малая РНК, продуцируемая вирусом в ходе этой программы. |
| miRNAs | латентные | микроРНК EBV кодируются двумя транскриптами, одним установлен в гене BART и один рядом с кластером BHRF1. Три BHRF1 pri-miRNAs (генерирующие четыре miRNA) экспрессируются во время латентного периода III типа, тогда как большой кластер miRNA BART (до 20 miRNA) экспрессируется во время латентного периода II типа. Функции этих miRNA в настоящее время неизвестны. |
| литический | ранний антиген | |
| литический | мембранный антиген | |
| EBV-VCA | литический | вирусный капсидный антиген |
| EBV-AN | литическая | щелочная нуклеаза |
EBV можно разделить на два основных типа: EBV типа 1 и EBV типа 2. Эти два подтипа имеют разные EBNA-3 гены. В результате два подтипа различаются по своим способностям к преобразованию и способности к реактивации. Тип 1 доминирует в большей части мира, но оба типа одинаково распространены в Африке. Можно отличить EBV типа 1 от EBV типа 2, разрезав вирусный геном с помощью рестрикционного фермента и сравнив полученные в результате образцы пищеварения с помощью гель-электрофореза.
EBV был замешан в нескольких заболеваниях, включая инфекционный мононуклеоз, лимфому Беркитта, лимфому Ходжкина, рак желудка, рак носоглотки, рассеянный склероз и лимфоматоидный гранулематоз. В частности, было показано, что B-клетки, инфицированные EBV, находятся в поражениях мозга пациентов с рассеянным склерозом. Дополнительные заболевания, которые были связаны с ВЭБ, включают синдром Джанотти – Крости, многоформную эритему, острые генитальные язвы, волосатую лейкоплакию полости рта. Вирусная инфекция также связана с широким спектром незлокачественных лимфопролиферативных заболеваний, таких как тяжелая гиперчувствительность аллергия на укус комара, Эпштейн, и часто способствует их развитию. -Положительные к вирусу Барра кожно-слизистые язвы и вакцина гидроа, а также злокачественные лимфопролиферативные заболевания, такие как лимфома Беркитта с положительной реакцией на вирус Эпштейна-Барра, вирус Эпштейна-Барра -положительная лимфома Ходжкина и первичная выпотная лимфома.
Вирус Эпштейна-Барра был вовлечен в нарушения, связанные с агрегацией альфа-синуклеина (например, болезнь Паркинсона, деменция с тельцами Леви и множественная системная атрофия ).
Вирус Эпштейна-Барра был назван в честь Майкла Энтони Эпштейна (родился 18 мая 1921), ныне почетный профессор Бристольского университета, и Ивонн Барр (1932–2016), 1966 доктор философии, выпускница Лондонский университет, которые вместе обнаружили и в 1964 году опубликовали информацию о существовании вируса. В 1961 году Эпштейн, патолог и эксперт электронный микроскоп, посетил лекцию на тему «Самый распространенный детский рак в тропической Африке - доселе нераспознанный синдром». Эта лекция Дениса Парсонс Беркитта, хирурга, практикующего в Уганде, была описанием «эндемичного варианта» (детской формы) болезни, которая носит его имя. В 1963 году образец был отправлен из Уганды в больницу Мидлсекса для культивирования. В культивируемых клетках были идентифицированы вирусные частицы, и результаты были опубликованы в The Lancet в 1964 году Epstein, Bert Achong и Barr. Линии клеток отправляли Вернеру и Гертруде Хенле в Детскую больницу Филадельфии, которые разработали серологические маркеры. В 1967 году техник в их лаборатории развил мононуклеоз, и они смогли сравнить сохраненный образец сыворотки, показав, что развиваются антитела к вирусу. В 1968 году они обнаружили, что EBV может напрямую иммортализовать B-клетки после заражения, имитируя некоторые формы EBV-инфекций, и подтвердили связь между вирусом и инфекционным мононуклеозом.
Как относительно сложный вирус, EBV еще полностью не изучен. Лаборатории по всему миру продолжают изучать вирус и разрабатывать новые способы лечения вызываемых им заболеваний. Одним из популярных способов изучения EBV in vitro является использование бактериальных искусственных хромосом. Вирус Эпштейна-Барра можно поддерживать и манипулировать в лаборатории в непрерывном латентном режиме (свойство, общее с вирусом герпеса Капоши, связанным с саркомой, другим из восьми вирусов герпеса человека). Хотя предполагается, что многие вирусы обладают этим свойством во время заражения их естественных хозяев, не существует легко управляемой системы для изучения этой части жизненного цикла вируса. Геномные исследования EBV позволили изучить литическую реактивацию и регуляцию латентного вирусного эписома.
Несмотря на активные исследования, вакцина против вируса Эпштейна-Барра еще не доступна. Разработка эффективной вакцины может предотвратить до 200 000 случаев рака во всем мире. Подобно другому вирусу герпеса человека 'Эпштейн-Барр может позволить эрадикацию с помощью курса пролекарства валацикловир, но необходимы дальнейшие исследования, чтобы определить, достижима ли эрадикация.