Войти
| ELK1 | |||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||
| |||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||
| Псевдонимы | ELK1, фактор транскрипции ETS, Фактор транскрипции ETS ELK1 | ||||||||||||||||||||||||||
| Внешние идентификаторы | OMIM: 311040 MGI: 101833 HomoloGene: 3832 Генные карты: ELK1 | ||||||||||||||||||||||||||
| |||||||||||||||||||||||||||
| |||||||||||||||||||||||||||
| |||||||||||||||||||||||||||
| Ортологи | |||||||||||||||||||||||||||
| Виды | Человек | Мышь | |||||||||||||||||||||||||
| Энтрез | |||||||||||||||||||||||||||
| Ensembl | |||||||||||||||||||||||||||
| UniProt | |||||||||||||||||||||||||||
| RefSeq (мРНК) |
ETS Like-1 белок Elk-1 - это белок, который у человека кодируется ELK1 . Elk-1 действует как активатор транскрипции. Он классифицируется как тройной комплексный фактор (TCF), подкласс семейства ETS, который характеризуется общим белковым доменом, который регулирует связывание ДНК с целевые последовательности. Elk1 играет важную роль в различных контекстах, включая формирование долговременной памяти, наркозависимость, болезнь Альцгеймера, синдром Дауна, рак груди и депрессия. Содержание
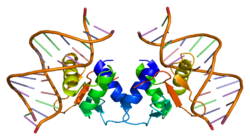
Структура  Рисунок 1 Рисунок 1 Как показано на рисунке 1, белок Elk1 состоит из нескольких доменов. Расположенный в N-концевой области, A-домен необходим для связывания Elk1 с ДНК. Эта область также содержит сигнал ядерной локализации (NLS) и сигнал ядерного экспорта (NES), которые отвечают за ядерный импорт и экспорт, соответственно. Домен B позволяет Elk1 связываться с димером его кофактора, фактор ответа сыворотки (SRF). Расположенный рядом с доменом B, домен R участвует в подавлении транскрипционной активности Elk1. Этот домен содержит остатки лизина, которые могут подвергаться SUMOylation, посттрансляционному событию, которое усиливает ингибирующую функцию R-домена. Домен D играет ключевую роль в связывании с активными митоген-активируемыми протеинкиназами (MAPK). Расположенный в C-концевой области Elk1, C домен включает аминокислоты, которые фактически становятся фосфорилированными с помощью MAPK. В этой области серин 383 и 389 являются ключевыми сайтами, которые необходимо фосфорилировать, чтобы могла произойти транскрипция, опосредованная Elk1. Наконец, домен DEF специфичен для взаимодействия активированной киназы, регулируемой внеклеточными сигналами (Erk), типа MAPK, с Elk1. Экспрессия Учитывая его играя роль фактора транскрипции, Elk1 экспрессируется в ядрах ненейрональных клеток. Белок присутствует в цитоплазме, а также в ядре зрелых нейронов. В постмитотических нейронах вариант Elk1, sElk1, экспрессируется только в ядре, поскольку он лишен сайта NES, присутствующего в полноразмерном белке. Более того, хотя Elk1 широко экспрессируется, фактические уровни варьируются в зависимости от тканей. Например, мозг крысы чрезвычайно богат Elk1, но белок экспрессируется исключительно в нейронах. Варианты сплайсинга Помимо полноразмерного белка, ген Elk1 может давать два сокращенные версии Elk1: ∆Elk1 и sElk1. Альтернативное соединение дает ∆Elk1. В этом варианте отсутствует часть ДНК-связывающего домена, которая позволяет взаимодействовать с SRF. С другой стороны, sElk1 имеет интактную область, которая связывается с SRF, но в нем отсутствуют первые 54 аминокислоты, содержащие NES. Обнаруженный только в нейронах, sElk1 создается с помощью внутреннего сайта начала трансляции. И ∆Elk1, и sElk1, усеченные версии полноразмерного белка, способны связываться с ДНК и индуцировать различные клеточные сигналы. Фактически, sElk1 противодействует Elk1 в дифференцировке нейронов и регуляции передачи сигналов фактора роста нервов / ERK. Передача сигналов Нисходящая мишень Elk1 является элементом ответа сыворотки (SRE) протоонкогена c-fos . Чтобы продуцировать c-fos, белок, кодируемый геном Fos, Elk1 должен быть фосфорилирован с помощью MAPK на его С-конце. MAPK являются последними эффекторами путей передачи сигнала, которые начинаются на плазматической мембране. Фосфорилирование с помощью MAPK приводит к конформационному изменению Elk1. Как видно на фиг. 2, киназа Raf действует выше MAPK, активируя их посредством фосфорилирования и, таким образом, активации MEK или киназ MAPK или ERK. Сам Raf активируется Ras, который связан с рецепторами фактора роста с активностью тирозинкиназы через Grb2 и Sos. Grb2 и Sos могут стимулировать Ras только после связывания факторов роста с их соответствующими рецепторами. Однако активация Raf не зависит исключительно от Ras. Протеинкиназа C, которая активируется сложными эфирами форбола, может выполнять ту же функцию, что и Ras. Киназа MEK (MEKK) также может активировать MEK, которые затем активируют MAPK, что иногда делает ненужным Raf. Следовательно, различные пути передачи сигнала проходят через MEK и MAPK и приводят к активации Elk1. После стимуляции Elk1 должен быть задействован SRF, который позволяет Elk1 связываться с промотором c-fos. Связывание Elk1 с SRF происходит из-за белок-белкового взаимодействия между B-доменом Elk1 и SRF и взаимодействия белок-ДНК через домен A. Вышеупомянутые белки похожи на рецепты для определенного сигнального выхода. Если один из этих ингредиентов, например SRF, отсутствует, выводится другой результат. В этом случае отсутствие SRF приводит к активации Elk1 другого гена. Таким образом, Elk1 может независимо взаимодействовать с сайтом связывания ETS, как в случае протоонкогена lck на рисунке 2. Более того, расстояние и относительная ориентация сайта связывания Elk1 с SRE довольно гибкие, что позволяет предположить, что SRE -регулируемые ранние гены, отличные от c-fos, могут быть мишенями для Elk1. egr-1 является примером цели Elk1, которая зависит от взаимодействия SRE. В конечном итоге фосфорилирование Elk1 может приводить к продукции многих белков, в зависимости от других вовлеченных факторов и их конкретных взаимодействий друг с другом. При изучении сигнальных путей мутации могут дополнительно подчеркнуть важность каждого компонента, используемого для активации нижестоящей мишени. Например, нарушение C-концевого домена Elk1, который MAPK фосфорилирует, запускает ингибирование активации c-fos. Сходным образом, дисфункциональный SRF, который обычно привязывает Elk1 к SRE, приводит к тому, что Fos не транскрибируется. В то же время без Elk1 SRF не может индуцировать транскрипцию c-fos после стимуляции MAPK. По этим причинам Elk1 представляет собой важное звено между путями передачи сигнала и инициацией транскрипции гена. Клиническая значимость Долговременная памятьФормирование долговременной памяти может зависеть от Elk1. Ингибиторы MEK блокируют фосфорилирование Elk1 и, таким образом, ухудшают приобретенное условное отвращение к вкусу. Более того, обучение избеганию, при котором субъект узнает, что конкретная реакция приводит к предотвращению аверсивного стимула, коррелирует с определенным увеличением активации Erk, Elk1 и c-fos в гиппокамп. Эта область мозга участвует в краткосрочном и долгосрочном хранении информации. Когда связывание Elk1 или SRF с ДНК блокируется в гиппокампе крысы, только секвестрация SRF нарушает долговременную пространственную память. Хотя взаимодействие Elk1 с ДНК может не быть существенным для формирования памяти, его специфическая роль все еще нуждается в изучении. Это связано с тем, что активация Elk1 может запускать другие молекулярные события, которые не требуют, чтобы Elk1 связывался с ДНК. Например, Elk1 участвует в фосфорилировании гистонов, усилении взаимодействия с SRF и рекрутировании основного транскрипционного аппарата, все из которых не требует прямого связывания Elk1 с ДНК. НаркоманияАктивация Elk1 играет центральную роль в наркозависимости. После того, как мышам дали кокаин, в полосатом теле наблюдается сильное и мгновенное гиперфосфорилирование Erk и Elk1. Когда этим мышам затем вводят ингибиторы MEK, фосфорилирование Elk1 отсутствует. Показано, что без активного Elk1 производство c-fos и вызванное кокаином обусловленное предпочтение места блокируются. Кроме того, острое употребление этанола приводит к чрезмерному фосфорилированию Elk1 в миндалине. Также было обнаружено, что подавление активности Elk1 снижает клеточные реакции на сигналы отмены и длительное лечение опиоидами, одним из старейших известных в мире препаратов. В целом эти результаты подчеркивают, что Elk1 является важным компонентом наркомании. ПатофизиологияПоказано, что накопление пептидов бета-амилоида (Aβ) вызывает и / или запускает Болезнь Альцгеймера. Aβ препятствует BDNF -индуцированному фосфорилированию Elk1. Поскольку активация Elk1 затруднена на этом пути, регуляция генов, управляемая SRE, приводит к повышенной уязвимости нейронов. Elk1 также ингибирует транскрипцию пресенилина 1 (PS1), который кодирует белок, необходимый для последней стадии последовательного протеолитического процессинга белка-предшественника амилоида (APP). APP создает варианты Aβ (полипептид Aβ42 / 43). Более того, PS1 генетически связан с большинством ранних случаев семейной болезни Альцгеймера. Эти данные подчеркивают интригующую связь между Aβ, Elk1 и PS1. Другое состояние, связанное с Elk1, - это синдром Дауна. У эмбриональных и старых мышей с этим патофизиологическим состоянием было обнаружено снижение активности кальциневрина, основной фосфатазы для Elk1. У этих мышей также есть возрастные изменения активации ERK. Более того, экспрессия SUMO3, который подавляет активность Elk1, увеличивается у взрослых пациентов с синдромом Дауна. Следовательно, синдром Дауна коррелирует с изменениями в путях ERK, кальциневрина и SUMO, все из которых действуют антагонистически на активность Elk1. Elk1 также взаимодействует с вариантами сплайсинга BRCA1, а именно BRCA1a и BRCA1b. Это взаимодействие усиливает опосредованное BRCA1 подавление роста клеток рака груди. Elk1 может быть нижестоящей мишенью BRCA1 в его пути контроля роста. Недавняя литература показывает, что активность промотора c-fos ингибируется, тогда как избыточная экспрессия BRCA1a / 1b снижает MEK-индуцированную активацию SRE. Эти результаты показывают, что один из механизмов подавления роста и опухоли белками BRCA1a / 1b действует посредством репрессии экспрессии последующих генов-мишеней Elk1, таких как Fos. Депрессия была связана с Elk1. Снижение Erk-опосредованного фосфорилирования Elk1 наблюдается в гиппокампе и префронтальной коре посмертного мозга суицидальных людей. Несбалансированная передача сигналов Erk коррелирует с депрессией и суицидальным поведением. Будущие исследования покажут точную роль Elk1 в патофизиологии депрессии. Ссылки Внешние ссылки
Последняя правка сделана 2021-05-18 14:55:23
Содержание доступно по лицензии CC BY-SA 3.0 (если не указано иное). | ||||||||||||||||||||||||||




 ..
..