
Войти
 12 популяций LTEE E. coli 25 июня 2008 г.
12 популяций LTEE E. coli 25 июня 2008 г. Модель E. coli долгосрочный эволюционный эксперимент (LTEE ) - это продолжающееся исследование экспериментальной эволюции под руководством Ричарда Ленски, которое используется отслеживало генетические изменения в 12 идентичных популяции бесполых бактерий Escherichia coli с 24 февраля 1988 г. Популяции достигли рубежа в 50 000 поколений в феврале 2010 г. 13 марта 2017 г. Ленски выполнил 10 000-й перенос эксперимента. Популяции достигли 73 500 поколений в начале 2020 года, незадолго до замораживания из-за пандемии COVID-19.
В ходе эксперимента Ленски и его коллеги сообщили о широком спектре фенотипических и генотипических изменений в наблюдениях. В их число вошли изменения, произошедшие во всех 12 популяциях, и другие, которые проявились в одном или нескольких популяциях. Например, во всех 12 популяциях наблюдалась схожая картина быстрое увеличение приспособления, которое со временем замедлялось, скорости роста и увеличения размера клеток. У половины популяций развились дефекты репарации ДНК, которые вызвали мутаторные фенотипы, отмеченные повышенным уровнем мутаций. Самая поразительная адаптация, о которой сообщалось до сих пор, - это эволюция аэробного роста на цитрате, что необычно для E. coli, в какой-то момент между поколениями 31000 и 31500.
Ленски принял решение приостановить эксперимент 9 марта 2020 года, когда его лаборатория временно закрылась в качестве меры предосторожности против распространения COVID19. Все линии длительного хранения были помещены в хранилище сверхнизких температур до тех пор, пока их можно разморозить для продолжения экспериментов.
4 мая 2020 года Ленски объявил о пятилетнем продлении гранта в рамках программы Долгосрочные исследования в области биологии окружающей среды (LTREB) национального научного фонда, которая поддерживает LTEE. Он также объявил, что эксперимент будет передан под наблюдение доктора Джеффри Э. Баррика, доцента кафедры молекулярных биологических наук Техасского университета в Остине, в течение следующих 5 лет. Доктор Баррик ранее был исследователем с докторской степенью у доктора Ленски и внес большой вклад в исследования, основанные на LTEE.
Долгосрочный эволюционный эксперимент был разработан как ограниченное средство эмпирического исследования центральных эволюций эволюции. Эксперимент был начат с тремя целями:
По мере продолжения эксперимента его масштабы росли, так как возникли новые вопросы в эволюционной биологии, которые могут использовать для решения, как эволюция популяций, представила новые явления для изучения, а также по мере развития технологий и методологических методов.
Использование E. coli в качестве экспериментального организма изучать многие поколения и большие популяции в относительно короткий период времени. Более того, из-за длительного использования E. coli в качестве основного модельного организма в молекулярной биологии, был доступен широкий спектр инструментов, протоколов и процедур для изучения изменений генетического, фенотипического и физиологического уровней.. Бактерии также можно заморозить и сохранить, оставаясь при этом жизнеспособными. Это позволяет создать то, что Ленски называет «замороженной летописью окаменелостей», состоящей из образцов эволюционирующих популяций, которые могут быть восстановлены в любое время. Эта замороженная летопись окаменелостей позволяет перезапускать образцы предков и других в эксперименте, а также позволяет изолировать и сравнивать живые образцы предков и эволюционировавших клонов. Ленски выбрал штамм E. coli, который воспроизводится только бесполым плазмиды, которые могли бы вызвать бактериальную конъюгацию, и не имеет жизнеспособного профага. Как следствие, эволюция в эксперименте происходит только за счет эволюционных процессов: мутации, генетического дрейфа и естественного отбора. Эта строгая асексуальность также означает, что генетические маркеры сохраняются в линиях и кладах по общему происхождению, но не иначе распространяться в популяциях.
Ленски решил провести эксперимент. с бактериями, выращенными в минимальной среде с ограничением глюкозы под названием DM25, которая была использована при выделении Бернардом Дэвисом для использования при выделении ауксотрофных мутантов E.coli с использованием пенициллина как селективный агент. DM25 содержит низкую концентрацию глюкозы. Ленски выбрал эту концентрацию, чтобы упростить анализ эволюции популяций за счет уменьшения уменьшения клональной интерференции, в которой несколько версий >Лелей конкурируют в эволюционирующей популяции, при этом уменьшая возможность эволюции. экологических взаимодействий. Эта используемой глюкозы поддерживает максимальную популяцию в 500 миллионов клеток предка в 10 мл культуры, хотя теперь максимально распространенных среди эволюциониров популяций. DM25 также содержит большое количество цитрата (примерно в 11 раз больше, чем глюкозы), который был включен Дэвисом, потому что он улучшил эффективность уничтожения пенициллина во время его экспериментов, хотя теперь известно, что он помогает в Получение E.coli железа из среды.
12 популяций в инкубаторе при 37 ° C (99 ° F) в лаборатории Ленски в Университета штата Мичиган. Каждый день 1% каждой популяции переносят в колбу со свежей питательной средой DM25. Разведение означает, что в каждой популяции происходит 6,64 поколения или удвоения. Большие репрезентативные образцы каждой популяции замораживают глицерином в качестве криопротектора с интервалами в 500 поколений (75 дней). Бактерии в этих образцах остаются жизнеспособными и могут быть восстановлены в любое время. Этот набор образцов называется «замороженной летописью окаменелостей» и обеспечивает эволюцию каждой популяции на протяжении всего эксперимента. Популяции также регулярно проверяются на предмет изменений в средней приспособленности. По состоянию на апрель 2016 г. популяции E. coli изучались на протяжении более 64 500 поколений, считая, что они претерпели достаточно спонтанных мутаций, чтобы все возможные одиночные точечные мутации в E. coli встречались несколько раз.
Штамм E. coli, который был получен из «штамма Bc251», как описано в статье 1966 года Сеймура Ледерберга через Брюса Левина, который использовал его в экспериментах по бактериальной экологии в 1972 году. Определяющими генетическими признаками этого штамма были: T6, Str, rm, Ara (не может расти на арабинозе ). Ленски обозначил оригинальный штамм-основатель как REL606. Перед началом эксперимента Ленски выделил Ara штамма, в котором вариант точечная мутация в опероне ara восстановила рост арабинозы, который он обозначил как штамм REL607. Долгосрочный эволюционный эксперимент, шесть поколений с шестью отдельными колониями Ara REL606. Эти популяции обозначают как Ara-1 - Ara-6. Ленский также основал еще шесть популяций из шести отдельных колоний ара REL607. Их называют популяциями от Ara + 1 до Ara + 6. Маркерные отличающиеся дифференцируемыми шмы на чашках Tetrazolium Arabinose, на которых колонии Ara + 1 красными, а колонии Ara выглядят от белых до розовых. В ходе эксперимента каждая популяция накопила большое количество различных мутаций, которые позволяют использовать дополнительные средства идентификации штаммов для популяции их происхождения.
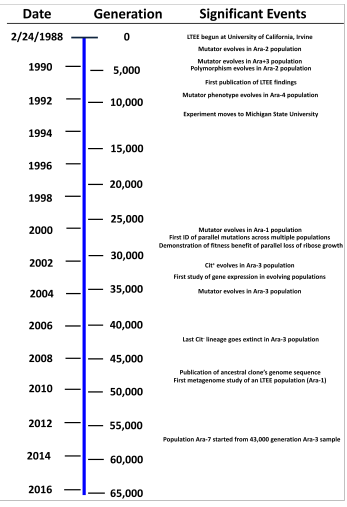 Хронология эксперимента по длительной эволюции E. coli, показывающая взаимосвязь между годами и поколениями эволюции, а также важные события и открытия.
Хронология эксперимента по длительной эволюции E. coli, показывающая взаимосвязь между годами и поколениями эволюции, а также важные события и открытия. Большой анализ эксперимента касался того, как изменилась приспособленность популяций по отношению к их предкам. Во всех популяциях наблюдалась периодерность увеличения относительной приспособленности в ранних ранних ранних ранних веков. К 20 000 поколениям популяция росла примерно на 70% быстрее, чем у предков. Это увеличение продолжалось в увеличении. Исследование 2013 Wiser et al. сообщили о постоянном улучшении на 50 000 поколений по с образцами, выделенными на 40 000 поколений. Они показывают, что увеличение приспособляемости намного лучше соответствует моделям степенного закона, чем гиперболические модели, которые использовались ранее. Как в популяризации фиксировались все более низкие полезные мутации, не превышающий верхний предел используется, как гиперболенная модель подразумевает предел, в работе, что будет продолжаться без ограничений. Дальнейшая работа, опубликованная в 2015 году, сообщила о результатах более 1100 новых тестов на пригодность, в которых изучались изменения приспособления на протяжении 60 000 поколений. Данные снова соответствуют предложенной модели степенного закона и действительно, соответствуют прогнозам модели на основе более ранних данных. Эти результаты предполагают, что, вопреки предыдущим представлениям, адаптация и адаптивная дивергенция могут увеличиваться бесконечно, даже в постоянных условиях.
Из 12 популяций на данный момент зарегистрировано шесть разрушения дефектов их возможностей восстанавливать ДНК, что значительно увеличило скорость мутаций в этих штаммах. Хотя используются бактерии в каждой популяции произвели миллионы мутаций в течение первых 20000 поколений, подсчитал, что в течение этого периода времени только от 10 до 20 полезных мутаций достигли фиксации в каждой популяции с менее 100 точечных мутаций (включая нейтральные мутации ), достигающих фиксации в каждой популяции. В 2009 году Barrick et al. Сообщает последовательность последовательностей генома из временных точек в поп-системе Ara-1. Они показывают, что, отличие от снижения скорости улучшения приспособленности, накопление мутаций было линейным и напоминало часы, хотя несколько линий показателей предполагали, что большая часть накопления была полезной, а не нейтральной.
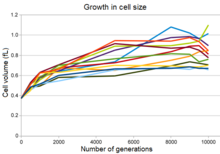 Рост клеток бактерий в эксперименте Ленски
Рост клеток бактерий в эксперименте Ленски Все двенадцать экспериментальных популяций увеличивают клетки одновременно с уменьшением максимальной плотности популяции, а во многих популяциях - более округлая форма ячеек. Это изменение было частично результатом мутации, которая изменила экспрессию гена пенициллин-связывающего белка, что позволяет мутантным бактериям вытеснить родовые бактерии в условиях длительного периода. -временной эволюционный эксперимент. Однако, хотя эта мутация повысила приспособленность в этих условиях, она также увеличила чувствительность бактерий к осмотический стрессу и снизила их способность выживать в течение длительного периода в культуре стационарной фазы.
В ходе экспериментов по производству специализированной продукции, в которой они используются глюкозы, на растут. Впервые это было описано в 2000 году, когда использовались физические функции после 20 000 поколений, что ограничило диапазон веществ, которые могли расти бактерии. Их анализ показал, что этот распад был вызван антагонистической плейотропией, при которой мутации улучшают способность расти на глюкозе, снижать или устранять способность расти на других веществах. Более используемое исследование Лейби и Маркса, в котором использовались более продвинутые методы, показало, что большая часть распада идентифицированного Купером и Ленски, была экспериментальными артефактами, что потеря неиспользуемых функций была не такой обширной, как предполагалось на первый взгляд. улучшены. Более, они пришли к выводу, что они были вызваны не антагонистической плейотропией, нейтральным накоплением мутаций в неиспользуемых частях генома, предполагая, что адаптация к простой среде не обязательно может привести к специализации.
Два различных варианта, S и L, идентифицированы в популяции обозначенной Ara-2, в 18000 поколений на основании их образования малых и больших колоний соответственно. Клоны типов S и L могли стабильно сосуществовать в совместном культивировании друг с другом, что указывает на то, что они занимаются разными ниши в популяции. Это было подтверждено открытием, L-тип преимущества во время роста глюкозе, но это было преимущество во время стационарной фазы после того, как глюкоза закончилась. Было обнаружено, что эти два типа развились до 6000 поколений, а затем сосуществовали после этого. Филогенетический анализ клонов двух типов, выделенных из разных поколений, показывает, что типы S и L принадлежат разным сосуществующим линиям в популяции и, возможно, претерпевают зарождающееся видообразование.
 Популяция, обозначенная Ara-3 (в центре), более мутная, потому что эта популяция эволюционировала, чтобы использовать цитрат, присутствующий в питательной среде.
Популяция, обозначенная Ara-3 (в центре), более мутная, потому что эта популяция эволюционировала, чтобы использовать цитрат, присутствующий в питательной среде. Э. coli обычно не может расти аэробно на цитрате из-за неспособности экспрессировать переносчик цитрата в присутствии кислорода. E. coli имеет полный цикл лимонной кислоты и, следовательно, метаболизирует цитрат в качестве промежуточного продукта во время аэробного роста других веществ, включая глюкозу. Большинство E. coli может расти анаэробно на цитрате посредством ферментации, если доступен дополнительный субстрат, такой как глюкоза, для обеспечения восстанавливающей способности. анаэробный рост возможен благодаря экспрессии трансмембранного гена цитрат-сукцинатного антипортера, citT, который был впервые идентифицирован в 1998 году. Этот ген совместно регулируется с другими генами, участвующими в цитратной ферментации, обнаруженными в цитрат-сукцинате. оперон, который включается только при отсутствии кислорода.
Неспособность к аэробному росту на цитрате, называемая фенотипом Cit, являющаяся определяющей характеристикой E. coli, имеющей значение ценным средством дифференциации кишечной палочки от патогенной сальмонеллы. Хотя штаммы Cit E. coli были выделены из образцов окружающей среды и сельскохозяйственных культур, в каждом таком случае было обнаружено, что этот признак связан с присутствием плазмиды, несущей чужеродный переносчик цитрата. О единственном спонтанном Cit мутанте E. coli сообщил Холл в 1982 году. Этот мутант был выделен во время длительного отбора для нового материала в среде питательной среды. Генетический анализ показал, что лежащая в основе мутации сложной, но в конечном итоге он смог идентифицировать точные изменения или задействованные гены, что привело его к гипотезе об активации загадочного гена-переносчика. Области генома, до Холлу, удалось сузить локализации изменений, не соответствуют прогнозному местоположению гена CitT, идентифицированному 16 лет спустя, а также физиологические характеристики аналитических мутантов Холла Cit не соответствуют прогнозируемым для аэробной экспрессия переносчика CitT.
В 2008 году команда Ленски во главе с Захари Д. Блаунтом сообщила, что способность к аэробному росту по цитрату развились в одной стимуляции. Примерно в поколении 33 127 наблюдалось резкое увеличение мутности в популяции, обозначенной как Ara-3. Они создали, что популяция содержала клоны, которые могли расти в аэробных условиях на цитрате (Cit). Эта способность способности популяции вырасти в несколько больше, чем раньше, из-за большого количества цитрата, присутствующего в среде. Изучение замороженных образцов ископаемых популяций показало, что клоны Cit могут быть выделены уже в 31 500 поколениях. Было обнаружено, что варианты Cit в популяции обладают рядом генетических маркеров, уникальных для популяции Ara-3; это исключало возможность того, что клоны были контаминантами, а не спонтанными мутантами. В экспериментах, «воспроизводили» ленту эволюции Ara-3 из клонов Cit, они использовали, что способность к аэробному росту на цитрате с большей вероятностью будет повторно развиваться в подмножество генетически чистых, эволюционирующих клонов. В этих экспериментах они наблюдали 19 новых случаев повторной эволюции Cit, когда они исходили из клонов, выделенных из более чем 20 000 поколений. Тесты флуктуации показали, что клоны этого поколения, а затем и более поздние показали скорость мутации по признаку Cit, которая была значительно выше, чем у предков. Даже в этих более поздних клонах скорость мутации одного порядка появления на триллион клеточных делений.
Ленски и его коллеги пришли к выводу, что эволюция функций Cit в этой одной популяции возникла из-за одного или более ранние, возможно, неадаптивные, «потенцирующие» мутации, которые увеличивают скорость мутации до доступного уровня. Данные свидетельствуют о том, что это связано как минимум с двумя мутациями после этих «усиливающих» мутаций. В более общем плане авторы предполагают, что историческая случайность может иметь глубокое и длительное влияние »на ходции, что указывает на следуя аргумент Стивена Джея Гулда. Эти открытия стали рассматриваться как важный пример исторической случайности на эволюцию.
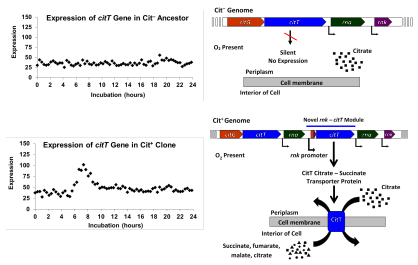 Признак Cit был актуализирован в результате дупликационной мутации, который создает новый регуляторный модуль путем помещения копии citT, кодирует цитрат-сукцинатный антипортер, под контролем поддерживает экспрессию в аэробных условиях. Эта мутация приводит к экспрессии переносчика CitT в присутствии кислорода, что позволяет расти на цитрате.
Признак Cit был актуализирован в результате дупликационной мутации, который создает новый регуляторный модуль путем помещения копии citT, кодирует цитрат-сукцинатный антипортер, под контролем поддерживает экспрессию в аэробных условиях. Эта мутация приводит к экспрессии переносчика CitT в присутствии кислорода, что позволяет расти на цитрате. В 2012 году Ленски и его команда сообщили о результатах геномного анализа признака Cit, проливающего свет на генетическую основу и историю эволюции. черты. Исследователи секвенировали полные геномы двадцати клонов, выделенные из различных точек в истории популяции Ara-3. Они использовали эти колонки, чтобы реконструировать филогенетическую историю популяции; Эта реконструкция показала, что население разделилось на три клады за 20 000 поколений. Варианты эволюционировали в одном из них, которые они назвали Clade 3. Клоны, потенцированные в более ранние исследования, были распределены между всеми тремя кладами, но были чрезмерно представлены в Clade 3. Это было по крайней мере две потенцирующие мутации., вовлеченные в эволюцию Cit.
Исследователи также представлены, что все клоны Cit имели мутации, в которых были сегмент ДНК из 2933 оснований был дуплицирован или амплифицирован. Дублированный сегмент содержания генов citT для белка-переносчика цитрата, используемого для анаэробного роста на цитрате. В результате этого используется лицом к другу по отношению друг к другу. Эта новая конфигурация помещает копию ранее молчащего, неэкспрессированного citT под контроль промотора соседнего гена rnk, который управляет экспрессией при наличии кислорода. Этот новый модуль rnk-citT создает новый регуляторный паттерн для citT, активируя экспрессию переносчика цитрата в кислород, и тем самым делает возможным аэробный рост цитрата.
Перемещение этого модуля rnk-citT в геноме показано, что усиленного клона Cit достаточно для получения фенотипа Cit. Однако исходный фенотип Cit, обусловленный дупликацией, был очень слабым и давал только ~ 1% -ное улучшение пригодности. Исследователи обнаружили, что количество копий модуля rnk-citT необходимо увеличить, чтобы усилить свойство цитрии в достаточной степени, чтобы бактерии хорошо выросли на цитрате. Дальнейшие мутации после того, как бактерии стали доминирующими в популяции, продолжали накапливать улучшенный рост цитрата.
Исследователи пришли к выводу, что эволюция признака Cit происходила в отдельные фазы: (1) накапливались мутации, которые увеличивали скорость мутации в Cit, (2) сам признак проявлялся в слабой форме и (3) признак был улучшен более поздними мутациями. Blount et al. Этот паттерн может быть типичным для эволюции новых черт в целом и трехэтапную модель эволюционных инноваций:
Эта модель получила признание в эволюционной биологии. В 2015 году палеонтолог Дуглас Эрвин использует модификацию четырехступенчатой модели, чтобы лучше отразить возможное различие между эволюционной новизной и функциональными инновациями, а также усиление условий окружающей среды: усиление, генерация новых фенотипов (актуализация), адаптивное усовершенствование и эксплуатация (преобразование новинки в нововведение, поскольку это становится важным для экологического становления организмов-обладателей).
В 2014 году исследовательская группа возглавила Эрик Квандт из лаборатории Джеффри Баррика в Техасском университете в Остине описал применение нового метода, называемого рекурсивной общегеномной рекомбинации и секвенирования (REGRES), для использования потенцирующих мутаций среди 70 присутствующих в арабском мире. 3 линия, которая эволюционировала Cit. В этом методе использовалось несколько раундов процесса, в котором плазмида F на основе конъюгации между клоном Cit 33000 поколения, CZB154, и клоном-основателем Cit LTEE для удаления мутаций, не требующихся ни для одного из них. проявление слабой или сильной формы Cit, которые они называют Cit. Они представляют, что модуль rnk-citT, ответственного за переключение фенотипа на Cit, было достаточно для создания слабого фенотипа Cit у предка. Они также идентифицировали мутацию, которая произошла в линии, ведущей к CZB154, которая предоставила сильный фенотип Cit у предка, отсутствовавший какой-либо мутацией, кроме модуля rnk-citT. Эта мутация, обнаруженная в регуляторной области гена, называемого dctA, вызвала массивное увеличение экспрессии транспортера DctA, которое выполняет функцию импорта C 4 -дикарбоксилатов в клетку. Они представляют, что эта повышенная экспрессия DctA позволяет CitT повторно поглощать сукцинат, малат и фумарат, высвобождаемые в среду транспортером CitT во время импорта цитрата. Они идентифицировали аналогичную мутацию в клонах Cit в популяции Ara-3, которая увеличивала экспрессию DctA за счет восстановления функций гена, который ее регулирует, dcuS, который был деактивирован в предковом клоне. Quandt et al. пришли к выводу, что мутация dctA участвует не в потенцировании, а в уточнении. Это эволюция в условиях генетической медицины. обеспечить значительное улучшение формы физической.
Квандт и его коллеги позже опубликовали результаты, окончательно идентифицирующие мутацию, которая действительно усиливала эволюцию Cit. Эта мутация была в гене gltA, который кодирует цитратсинтазу, фермент, участвующий в потоке углерода в цикл лимонной кислоты. Это имело эффект увеличения активности цитрат-синтазы, и они показали, что он позволяет улучшить рост ацетата. Более того, с мутацией gltA модуль rnk-citT, который включает признак Cit, имеет нейтральный или незначительно благоприятный эффект приспособления, тогда как без него модуль был бы сильно вредным. Следовательно, мутация gltA, по-видимому, позволяет ранним, слабым вариантам Cit сохраняться в популяции до тех пор, пока не могли произойти более поздние уточняющие мутации, в соответствии с более ранними выводами. После развития сильного фенотипа Cit повышенная активность цитратсинтазы стала пагубной. Исследователи обнаружили, что более поздние мутации в gltA противодействуют первой мутации, снижающей активность цитрат-синтазы и улучшая рост цитрата. Они пришли к выводу, что серия мутаций в gltA сначала усиливала, а затем улучшала рост цитрата. Они также предположили, что линия, в которой произошел Cit, может использовать производные Ara-3 для использования ацетата, и что потенцирующие мутации, которые приводят к эволюции Cit в Ara-3, изначально были адаптивными для использования ацетата.
Небольшая субпопуляция Cit-клеток, неспособных к росту цитратов и принадлежащих к ним кладе, сохранялась в популяции после того, как Cit-клетки стали доминирующими. Ранние результаты показывают, что это разнообразие частично объясняется тем, что Cit-клетки лучше растут на глюкозе в среде. Тернер и др. позже, клетки Cit развили способность перекрестно питаться большинством Cit. Они показали, что клетки Cit высвобождают сукцинат, малат и фумарат во время роста цитрате, поскольку переносчик CitT выкачивает эти вещества из клеток, перекачивая цитрат. в камеру. Клетки Cit быстро развили способность расти на этих веществах из-за мутации, которая восстановила экспрессию белка-переносчика, который молчал у предка.
Субпопуляция Cit в конечном итоге вымерла среди от 43 500 до 44 000 поколений. Было показано, что это вымирание не произошло из-за того, что большинство граждан Китая эволюционировало, чтобы иметь возможность вторгнуться в нишу, занятую меньшинством. Действительно, клоны Cit могут вторгаться в популяцию Cit после вымирания. «Более того, в устройстве, в котором перезапустили эксперимент повторения популяции Ara-3 из образца, замороженного за 500 поколений до исчезновения», Тернер и другие. присутствует, что субпопуляция Cit не вымерла ни в одном из повторений после 500 поколений эволюции. Одна из этих копий продолжалась 2500 поколений, в течение которых Cit продолжал сосуществовать. Исследователи пришли к выводу, что исчезновение Citi произошло из-за неизвестного «редкого нарушения окружающей среды», аналогичного тому, что может повлиять на естественные популяции. Последний экземпляр был интегрирован в основной эксперимент LTEE, став тринадцатой популяцией, Ara-7.
Другие исследователи экспериментировали с эволюцией аэробных цитрат-использующих E. coli. Дастин Ван Хофвеген и др., Работающие в лаборатории разумного замысла сторонника сторонника Скотта Минниха, применили 46 независимых цитратных мутантов E. coli всего за 12-100 поколений. используя очень продолжительный отбор при голодании, во время которого бактерии будут быстрее обнаруживать больше мутаций. В их исследовании секвенирование геномной ДНК выявлено усиление локусов citT и dctA, а перестройка ДНК была тем же классом мутаций, который был идентифицирован в эксперименте Ричарда Ленски и его команды. Они пришли к выводу, что редкость мутанта, использующего цитрат, в исследованиях Ленски, скорее всего, был результатом селективных экспериментальных условий, использу его командой, не уникальным образом эволюционного видообразования.
Джон Рот и Софи Майснье-Патен сделали обзор подходов к отложенным мутациям группы Ленски, так и к быстрым мутациям группы Ван Хофвегеса на E. coli. Они утверждают, что обе команды испытали одну и ту же последовательную последовательность, актуализации и уточнения, приведенную к аналогичным вариантам Cit. По их словам, период менее суток, в течение которого цитрата будет выбираться, следует 100-кратное разбавление и период роста глюкозы, которое не будет выбрана для использования цитрата, в конечном итоге снизив вероятность заражения E. coli. способность накапливать ранние адаптивные мутации от одного периода отбора к другому. С другой стороны, группа Ван Хофвегена наблюдела непрерывный период отбора продолжительностью 7 дней, что привело к более быстрому развитию E. coli, использующей цитрат. Пот и Майнье-Патен обеспечивают, что серийное разведение E. coli и короткий период отбора для использования цитрата в условиях LTEE постоянно препятствуют достижению каждого поколения E. coli на следующих стадиях аэробного использования цитрата.
В ответ Блаунт и Ленски признают, что проблема не в экспериментах или данных, а в интерпретациях, сделанных Ван Хофвегеном и др. и Мезнье-Патен и Рот. Ленски отмечает, что быстрая эволюция Cit не обязательно была неожиданностью, поскольку его команда также смогла произвести несколько мутантов Cit за несколько недель во время экспериментов по воспроизведению, о которых они сообщили в статье 2008 года, в которой его команда впервые описала эволюцию аэробного использования цитрата в LTEE. Кроме того, Ленски критикует описание Ван Хофвегена и др. Первоначальной эволюции Cit как «события видообразования», указывая, что LTEE не был разработан для выделения мутантов, использующих цитрат, или для борьбы с видообразованием, поскольку в своей статье 2008 года они сказал, что «превращение в Cit было только первым шагом на пути к возможному видообразованию», и, таким образом, не предполагал, что мутанты Cit были другим видом, но что видообразование могло быть конечным следствием эволюции признака. Ленски признает, что ученые, в том числе он и его команда, часто используют сокращенный язык и жаргон при обсуждении видообразования вместо того, чтобы писать более тщательно и точно по этому вопросу, и это может вызвать проблемы. Однако он отмечает, что видообразование обычно рассматривается биологами-эволюционистами как процесс, а не как событие. Он также критикует Ван Хофвегена и др. и Рот и Майснье-Патен за постулирование «ложных дихотомий» в отношении сложной концепции исторической случайности. Он утверждает, что историческая случайность означает, что история имеет значение, и что в их статье 2008 года представлены данные, показывающие, что эволюция Cit в LTEE зависела от мутаций, которые накапливались ранее. Он заключает, что «... историческая случайность была задействована и продемонстрирована в конкретном контексте, а именно в контексте появления Cit в LTEE - это не означает, что появление Cit исторически обусловлено в других экспериментальных контекстах, и в этом отношении что другие изменения в LTEE исторически обусловлены - на самом деле, некоторые другие эволюционные изменения в LTEE были в высшей степени предсказуемыми и не (или, по крайней мере, не очевидно) зависели от предыдущих мутаций в популяциях ».
| Викискладе есть медиафайлы, связанные с E. coli долгосрочный эволюционный эксперимент. |