
Войти
| Гаплогруппа E-V68 | |
|---|---|
| Возможное время происхождения | c.24000 лет назад |
| Возраст слияния | c.19 900 лет назад |
| Возможное место происхождения | Египет / Ливия или южный Египет / север Судан |
| Предок | E-M215-M35 |
| Потомки | E-M78, E-V1039 |
| Определение мутаций | V68, L539, PF2203 |
Гаплогруппа E-V68, также известная как E1b1b1a, является гаплогруппой ДНК Y-хромосомы человека, обнаруженной в Северной Африке, Африканской основной Роге, зарегистрированной и Европе. Это субклад более крупной и старой гаплогруппы, известной как E1b1b или E-M215 (также примерно эквивалент E-M35). Линия E1b1b1a идентифицируется по наличию мутации однонуклеотидного полиморфизма (SNP) на Y-хромосоме, которая известна как V68. Это предмет обсуждения и изучения в генетике, а также в генетической генеалогии, археологии и исторической лингвистике.
E-V68. его более субстанкладом E-M78. В различных публикациях и E-V68, и E-M78 упоминаются под другими названиями, особенно филогенетической номенклатурой, такой как «E3b1a», которые предназначены для обозначения их места в генеалогическом древе всех мужчин-мужчин. Эти различные названия меняются по мере новых открытий и обсуждаются ниже.
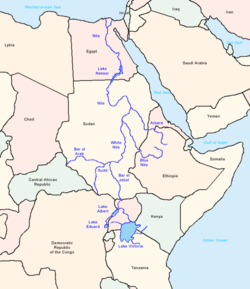 Река Нил и ее основные притоки: вероятный коридор миграций древних людей, включая те, которые связаны с линиями ДНК Y E-M243, E-M78, E-V12 и E-V22.
Река Нил и ее основные притоки: вероятный коридор миграций древних людей, включая те, которые связаны с линиями ДНК Y E-M243, E-M78, E-V12 и E-V22.E-M78, как и его родительская клада E-V68, как полагают, Afr Иканское происхождение. Основываясь на данных генетической вариации STR, Cruciani et al. (2007) предполагает, что этот субклад возник в «Северо-Восточной Африке», что в исследовании относится к региону Египта и Ливии.
До Cruciani et al.. (2007), Semino et al. (2004) интересное место происхождения E-M78 дальше на юг в Восточной Африке. Это произошло из-за высокой частоты и разнообразия линий E-M78 в регионе Эфиопия. Однако Cruciani et al. (2007) смогли изучить больше данных и пришли к выводу, что в линии E-M78 в Африканском Роге преобладали относительно недавние ветви (см. E-V32 ниже). Они пришли к выводу, что регион Египта был вероятным местом происхождения E-M78, сфера деятельности на периферийном географическом распределении наиболее производных подгаплогрупп по отношению к северо-востоку от Африки, а также результаты количественного анализа UEP и микросателлитного разнообразия ».
Cruciani et al. (2007) также отмечают это как свидетельство «коридора для двунаправленных миграций» между Северо-Восточной Африкой (Египет и Ливия по их данным) с одной стороны и Восточной Африки с другой. <Времена 171>Cruciani et al. (2007) также предположили, что E-M35, родительский класс E-M78, возник в Восточной Африке во время палеолита и распространился в регионе Египта. Таким образом, E-M78 в Восточной Африке является альтернативным движением. Авторы считают, что было «как минимум 2 эпизода между 23,9–17,3 тыс. Руб. Лет назад и 18,0–5,9 тыс. Руб. Лет назад ».
Другая вероятная миграция на юг из Египта была отмечена Hassan et al. (2008) на основе их исследования Судана. В частности, E-V12 и E-V22 «могли быть привезены в Судан из Северной Африки после прогрессирующего опустынивания Сахары около 6000-8000 лет назад».
К северу от Египта и Ливии E-M78 мигрировал на Ближний Восток, но, кроме того, Trombetta et al. (2011) предположили, что более ранняя популяция, несущая E-V68, могла мигрировать морем непосредственно из Африки в юго-западную Европу, поскольку они наблюдали случаи E-V68 * (без мутации M78) только в Сардинии И не в ближневосточных образцах. Что касается E-M78, как и других форм E-V68, есть свидетельства множественных путей экспансии за пределы африканской родины.
С другой стороны, хотя очевидно, что имели место перевозки из Северной Африки в Иберию и Южную Италию (людей, несущих E-V68 *, E-V12, E-V22 и E-V65), большинство линий E-M78, обнаруженных в Европе, принадлежащих к субкладу E-V13, который, по-предположительно, проник в Европу в какое-то время, не определенное с Ближнего Востока, где он очевидно, возникла через Балканы.
Придя к таким же выводам, что и команда Круциани и Тромбетта, Батталья и др. (2008), писавшие до открытия E-V68, описывают Египет как «центр распространения различных географически локализованных подкладов, связанных с M78», и, они предполагают, что точка происхождения E-M78 (в отличие от более поздних расселений из Египта), возможно, находился в , который «существовал на границе современного Судана и Египта, около Озеро Нубия, до начала влажной фазы около 8500 г. до н. Э. Движущиеся на север дождевых пояса в этот период могут стимулировать миграцию мезолитических собирателей на север в Африке, Левант и, в конечном итоге, до Малой Азии и Европы, где каждый в конечном итоге разделился на свои региональные отличительные ветви ".
Разделение E-V68 на подклассы, такие как E-V12, E-V13 и т. Д., В степени было делом работы итальянской команды Они начали на основе исследований STR в 2004 году, а в 2006 году они объявили об открытии мутаций однонуклеотидного полиморфизма (SNP), включая Фульвио Кручиани, Бениамино Тромбетта, Розарио Скоццари и других. Эти статьи были обновленными филогениями, найденными в Karafet (2008) harvcoltxt error: нет цели: CITEREFKarafet2008 (help ), и ISOGG, который, в свою очередь, является ос новой приведенной ниже филогении.
Loosdrecht et al. (2018) проанализировали полногеномные данные семи древних иберомавров особей из Grotte des Pigeons Taforalt в восточном около Марокко. Окаменелости были непосредственно датированы периодом между 15 100 и 13 900 калиброванными годами до настоящего времени. Ученые обнаружили, что все мужские особи с достаточной сохранностью ядерной ДНК принадлежали к субкладу E1b1b1a1 (M78), причем один скелет нес родительскую линию E1b1b1a1b1 к E-V13.
Батталья и др. (2007) ошибка harvcoltxt: нет цели: CITEREFBattagliaFornarinoAl-ZaheryOlivieri2007 (help ) подсчитал, что E-M78 (в этой статье называемый E1b1b1a1) находится в Европе более 10 000 лет. А совсем недавно Лакан и др. (2011) появились, что человеческие останки, раскопанные в испанской погребальной пещере, датируемые примерно 7000 лет назад, находились в ветви E-V13 E-M78.
В июне 2015 года мутация M78 и последующее начало генеалогических деревьев E-M78 и E-V68 были датированы Trombetta et al. примерно 20 300-14 800 лет назад.
Это филогенетическое дерево субкладов гаплогрупп основано на дереве ISOGG 2019.
| V68 | E-V68 * (E1b1b1a *) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| M78 |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
На данный момент на Сардинии Trombetta et al. Зарегистрированы три человека, которые находятся в E-V68, но не в E-M78. (2010) ошибка harvcoltxt: нет цели: CITEREFTrombettaCrucianiSellittoScozzari2010 (help ) при объявлении об открытии V68.
E-M78 широко распространен в Северной Африке, Африканском Роге, Западной Азии (простираясь до Южной Азии ) и Европа.
Самая базовая и редкая парагруппа E-M78 * была обнаружена с самыми высокими частотами у египтян из оазиса Гурна (5,88%), более низкие частоты также наблюдаются у марокканских арабов, сардинцев, Балкан и андалузцев из Уэльва.
Самые высокие частоты определенных подклассов E-M78 в основном обнаруживаются афроазиатских говорящих популяций на большой территории, простирающейся от предполагаемого места происхождения гапруппы в Верхнем Египте до Судан и Африканский Рог.
За пределами этой области распространения (Северная Африка и Африканский Рог) E-V68 также присутствует в других частях континента в более низких частотах из-за более поздних разгонов. Таким образом, сегодня он обнаружен в карманах Великих африканских озер и Южной Африки благодаря ранним афро-азиатским поселенцам из региона Горн и на западе, до Гвинеи. -Бисау, где его присутствие было приписано транссахарским перемещением людей из Северной Африки.
Распространение E-V68 в Европе определяет его субкладом E-V13, за исключением Иберии.. E-V13 имеет пик частоты с центром в некоторых частях Балкан (примерно 20% в южных районах; до почти 50% - в некоторых местах и населенных пунктах). Сегодня он имеет более низкие частоты в западных, центральных и северо-восточных регионах, хотя E-V13 был обнаружен в захоронении эпохи неолита в Каталонии. Это обсуждается более подробно ниже.
| Регион | Население | n | E-M78 | E-M78* | E-V12* | E-V13 | E-V22 | E-V32 | E-V65 | Исследование |
|---|---|---|---|---|---|---|---|---|---|---|
| Европа | албанцы | 55 | 25,46% = (14/55) | 1,82% = (1/55) | 23,64% = (13/55) | |||||
| Европа | македонские албанцы | 64 | 35,94% = (23 / 64) | 1,56% = (1/64) | 34,38% = (22/64) | |||||
| Европа | албанцы +. македонские албанцы | 55+. 64 =. 119 | 31,09% = (37/119) | 1,68% = (2/119) | 29,41% = (35/119) | |||||
| Европа | косовские албанцы | 114 | 45,61% = (52/114) | 1,75% = (2/114) | 43,86% = (50/114) | Peričic et al. (2005) ошибка harvcoltxt: нет цели: CITEREFPeričic_et_al.2005 (help ) | ||||
| Европа | албанцы | 96 | 32,29% = (31/96) | 32,29% = ( 31/96) | Cruciani et al. (2007) | |||||
| Европа | косовские албанцы +. македонские албанцы +. албанцы | 119+. 114+. 96 =. 329 | 36,47% = (120/329) | 1,22% = (4/329) | 35,26% = (116/329) | .. Peričic et al. (2005) ошибка harvcoltxt: нет цели: CITEREFPeričic_et_al.2005 (help ). Cruciani et al. (2007) | ||||
| Европа | македонские арумана | 57 | 29,82 | 29, 82 | Peričic et al. (2005) ошибка harvcoltxt: нет цели: CITEREFPeričic_et_al.2005 (help ) | |||||
| Europe | сербы | 113 | 20,35 | 1,77 | 18,58 | Peričic et al. (2005) harvcoltxt error: no target: CITEREFPeričic_et_al.2005 (help ) | ||||
| Европа | хорваты | 108 | 5.60 | 5.60 | Peričic et al. (2005) ошибка harvcoltxt: нет цели: CITEREFPeričic_et_al.2005 (помощь ) | |||||
| Европа | Крит | 1 93 | 6,7% = 13/193 | 6,7% = 13/193 | King et al. (2008) ошибка harvcoltxt: нет цели: CITEREFKing_et_al.2008 (help ) | |||||
| Europe | Греки из Неа Никомедеи | 57 | 15,8% = 9/57 | 1,8% = 1/57 | 14,0% = 8/57 | King et al. (2008) ошибка harvcoltxt: нет цели: CITEREFKing_et_al.2008 (help ) | ||||
| Европа | Греки из Сескло / Димини | 57 | 38,6% = 22/57 | 3,5% = 2/57 | 35,1% = 20/57 | Кинг и др. (2008) harvcoltxt ошибка: нет цели: CITEREFKing_et_al.2008 (help ) | ||||
| Europe | Греки из Лерны / Franchthi | 57 | 35,1% = 20 / 57 | 35,1% = 20/57 | King и др. (2008) ошибка harvcoltxt: нет цели: CITEREFKing_et_al.2008 (help ) | |||||
| Европа | Греки с Крита +. греки из Неа Никомедеи. Греки из Сескло / Димини. из Лерны / Франчти | 193+. 57+. 57+. 57 =. 364 | 17, 58% = 64/364 | 0,82% = 3/364 | 16,76% = 61/364 | Кинг и др. (2008) ошибка harvcoltxt: нет цели: CITEREFKing_et_al. 2008 (help ) | ||||
| Европа | Континентальные греки | 147 | 19,05% = 28/147 | 17,69% = 26/147 | 0,68% = 1/147 | 0,68% = 1/147 | Cruciani et al. (2007) | |||
| Европа | Греки с Крита | 215 | 6,51% = 14/215 | 0,93% = 2/215 | 5,58% = 12/215 | Cruciani et al. (2007) | ||||
| Европа | Греки с Эгейских островов | 71 | 16,9% = 12/71 | 15,49% = 11/71 | 1,41% = 1/71 | Cruciani et al. (2007) | ||||
| Европа | Континентальные греки. Греки с Крита. Греки с Эгейских островов | 147+. 215+. 71 =. 433 | 12,47 % = 54/433 | 0,46% = 2/433 | 11,32% = 49/433 | 0,46% = 2/433 | 0,23% = 1/433 | Cruciani et al. (2007) | ||
| Европа | Греки с Крита +. Греки из Неа Никомедеи. Греки из Сескло / Димини. из Лерны / Франхти. Континентальные греки. Греки с Крита. Греки с Эгейских островов | 364+. 433 =. 797 | 14,81% = 118/797 | 0,38% = 3/797 | 0,25% = 2 / 797 | 13,8% = 110/797 | 0,25% = 2/797 | 0,13% = 1/797 | King et al. (2008) ошибка harvcoltxt: нет цели: CITEREFKing_et_al.2008 (help ). Cruciani et al. (2007) | |
| Европа | сицилийцы | 236 | 11,43 | 1,27 | 5,93 | 3,81 | 0,42 | Ди Гаэтано и др. (2008) ошибка harvcoltxt: нет цели: CITEREFDi_Gaetano_et_al.2008 (помощь ) | ||
| Европа | Андалузцы Уэльвы | 167 | 6,59 | 1,20 | 4,19 | 0,60 | 0,60 | Ambrosio et al. (2010) ошибка harvcoltxt: нет цели: CITEREFAmbrosio_et_al.2010 (help ) | ||
| Европа | македонцы | 99 | 18,18 | 17,17 | 1.01 | Cruciani et al. (2007) | ||||
| Европа | Болгары | 204 | 16,67 | 0,49 | 16,18 | Cruciani et al. (2007) | ||||
| Европа | сицилийцы | 153 | 13,07 | 0,65 | 7,19 | 4,58 | 0,65 | Cruciani et al. (2007) | ||
| Европа | северные итальянцы | 94 | 7,45 | 5,32 | 2,13 | Cruciani и др. ( 2007) | ||||
| Европа | Центральные итальянцы | 356 | 7,87 | 0,28 | 5,34 | 1, 97 | 0,28 | Cruciani et al. (2007) | ||
| Европа | Южные итальянцы | 141 | 10,64 | 0,71 | 8,51 | 1,42 | Cruciani et al. (2007) | |||
| Европа | Сардинцы | 374 | 3,48 | 0,27 | 0,27 | 1,07 | 0,8 | 1,07 | Cruciani et al. (2007) | |
| Европа | Северный Португальский | 50 | 4 | 4 | Cruciani et al. (2007) | |||||
| Европа | Южный Португальский | 49 | 4,08 | 4,08 | Cruciani et al. (2007) | |||||
| Европа | Пасиегос из Кантабрии | 56 | Cruciani et al. (2007) | |||||||
| Европа | Астурийцы | 90 | 10 | 5,56 | 4,44 | Cruciani et al. (2007) | ||||
| Европа | Южные испанцы | 62 | 3,23 | 3,23 | Cruciani et al. (2007) | |||||
| Европа | Испанские баски | 55 | Cruciani et al. (2007) | |||||||
| Европа | Французские баски | 16 | 6,25 | 6,25 | Cruciani et al. (2007) | |||||
| Европа | Французский | 225 | 4,44 | 0,44 | 4 | Cruciani et al. (2007) | ||||
| Европа | английский | 28 | Cruciani et al. (2007) | |||||||
| Европа | Датский | 35 | 2,86 | 2,86 | Cruciani et al. (2007) | |||||
| Европа | Немцы | 77 | 3,9 | 3,9 | Cruciani et al. (2007) | |||||
| Европа | Польский | 40 | 2,5 | 2,5 | Cruciani et al. (2007) | |||||
| Европа | Чехи | 268 | 4,85 | 4,85 | Cruciani et al. (2007) | |||||
| Европа | Словаки | 24 | 8,33 | 8,33 | Cruciani et al. (2007) | |||||
| Европа | Словенцы | 104 | 2.88 | 2.88 | Cruciani et al. (2007) | |||||
| Европа | Эстонцы | 74 | 4,05 | 4,05 | Cruciani et al. (2007) | |||||
| Европа | белорусы | 40 | Cruciani et al. (2007) | |||||||
| Европа | Северные русские | 82 | 3,66 | 3,66 | Cruciani et al. (2007) | |||||
| Европа | Южные русские | 92 | 2,17 | 2,17 | Cruciani et al. (2007) | |||||
| Европа | Украинцы | 11 | 9.09 | 9.09 | Cruciani et al. (2007) | |||||
| Европа | Молдаване | 77 | 7,79 | 7,79 | Cruciani et al. (2007) | |||||
| Европа | Венгры | 106 | 9,43 | 9,43 | Cruciani et al. (2007) | |||||
| Европа | Румыны | 265 | 7,55 | 7,17 | 0,38 | Cruciani et al. al. (2007) | ||||
| Северо-Западная Африка | Марокканские арабы | 55 | 40 | 3,64 | 7,27 | 29,09 | Cruciani et al. (2007) | |||
| Северо-Западная Африка | Асни Берберс | 54 | 3,7 | 3,7 | Cruciani et al. (2007) | |||||
| Северо-Западная Африка | Бухрия Берберс | 67 | 1,49 | 1,49 | Cruciani et al. (2007) | |||||
| Северо-Западная Африка | Moyen Atlas Berbers | 69 | 10,14 | 10,14 | Cruciani et al. (2007) | |||||
| Северо-Западная Африка | Марракешские берберы | 29 | 6,9 | 3,45 | 3,45 | Cruciani et al. (2007) | ||||
| Северо-Западная Африка | Марокканские евреи | 50 | 12 | 2 | 2 | 8 | Cruciani et al. (2007) | |||
| Северо-Западная Африка | Мозабитские берберы | 20 | Cruciani et al. (2007) | |||||||
| Северо-Восточная Африка | Ливийские евреи | 25 | 8 | 4 | 4 | Cruciani et al. (2007) | ||||
| Северо-Восточная Африка | Ливийские арабы | 10 | 20 | 20 | Cruciani et al. (2007) | |||||
| Северо-Восточная Африка | Северные египтяне (Дельта) | 72 | 23,61 | 5,56 | 1,39 | 13, 89 | 2,78 | Cruciani et al. (2007) | ||
| Северо-Восточная Африка | Египетские берберы | 93 | 6,45 | 2,15 | 4,3 | Cruciani et al. (2007) | ||||
| Северо-Восточная Африка | Египтяне из Бахари | 41 | 41,46 | 14,63 | 2,44 | 21,95 | 2,44 | Cruciani et al. (2007) | ||
| Северо-Восточная Африка | Египтяне из оазиса Гурна | 34 | 17,65 | 5,88 | 8,82 | 2,94 | Cruciani et al. (2007) | |||
| Северо-Восточная Африка | Египтяне | 70 | 79 | 79 | Тромбетта (2015) | |||||
| Северо-Восточная Африка | Южные египтяне | 79 | 50,63 | 44, 3 | 1,27 | 3,8 | 1,27 | Cruciani et al. (2007) | ||
| Восточная Африка | Динка | 26 | 15,38 | 3,85 | 11,54 | Hassan et al. (2008) | ||||
| Восточная Африка | Шиллук | 15 | 13,33 | 13,33 | Hassan et al. (2008) | |||||
| Восточная Африка | Nuer | 12 | 16,67 | 16,67 | Hassan et al. (2008) | |||||
| Восточная Африка | Боргу | 26 | 15,38 | 3,85 | 11,54 | Hassan et al. (2008) | ||||
| Восточная Африка | Нуба | 28 | 25 | 3,57 | 3,57 | 7,14 | 10,71 | Hassan et al. al. (2008) | ||
| Восточная Африка | Масалит | 32 | 71,88 | 3,13 | 15,63 | 53,13 | Hassan et al. al. (2008) | |||
| Восточная Африка | Мех | 32 | 59,38 | 18,75 | 40,63 | Hassan et al. (2008) | ||||
| Восточная Африка | нубийцы | 39 | 15,38 | 12,82 | 2,56 | Hassan et al. (2008) | ||||
| Восточная Африка | Фулани из Судана | 26 | 34,62 | 30,77 | 3,85 | Hassan et al. (2008) | ||||
| Восточная Африка | Хауса из Судана | 32 | 3,13 | 3,13 | Hassan et al. (2008) | |||||
| Восточная Африка | Египетские копты из Судана | 33 | 15,15 | 15,15 | Hassan et al. (2008) | |||||
| Восточная Африка | Бежа | 42 | 35,71 | 4,76 | 30,95 | Hassan et al. (2008) | ||||
| Восточная Африка | Gaalien | 50 | 18,00 | 6,00 | 6,00 | 6,00 | Hassan et al. (2008) | |||
| Восточная Африка | Meseria | 28 | 14,29 | 3,57 | 10,71 | Hassan et al. (2008) | ||||
| Восточная Африка | Аракиен | 24 | 16,67 | 8,33 | 4,17 | 4,17 | Hassan et al. (2008) | |||
| Восточная Африка | Амхара | 34 | 8,82 | 8,82 | Cruciani et al. (2007) | |||||
| Восточная Африка | Эфиопские евреи | 22 | 9,09 | 9,09 | Cruciani et al. (2007) | |||||
| Восточная Африка | Смешанные эфиопы | 12 | 33,33 | 25 | 8,33 | Cruciani et al. (2007) | ||||
| Восточная Африка | Борана / Оромо (Кения / Эфиопия) | 32 | 40,63 | 40,63 | Cruciani et al. (2007) | |||||
| Восточная Африка | Волайта | 12 | 16,67 | 8,33 | 8,33 | Cruciani et al. (2007) | ||||
| Восточная Африка | Сахо из Эритреи | 94 | 88,3 | 88,3 | Тромбетта (2015) | |||||
| Восточная Африка | Сомали из Эфиопии | 12 | 33,3 | 8,3 | 25 | Тромбетта (2015) | ||||
| Восточная Африка | Сомали из Сомали | 5 | 80 | 80 | Тромбетта (2015) | |||||
| Восточная Африка | Сомали из Кении | 6 | 80 | 80 | Тромбетта (2015) | |||||
| Восточная Африка | Нилотик из Кении | 18 | 11,11 | 11,11 | Cruciani et al. (2007) | |||||
| Восточная Африка | банту из Кении | 28 | 3,57 | 3,57 | Cruciani et al. (2007) | |||||
| Восточная Африка | Западная Африка | 123 | 0,81 | 0,81 | Cruciani et al. (2007) | |||||
| Восточная Африка | Центральная Африка | 150 | 0,67 | 0,67 | Cruciani et al. (2007) | |||||
| Восточная Африка | Южная Африка | 105 | Cruciani et al. (2007) | |||||||
| Западная Азия | Стамбул Турецкий | 35 | 8,57 | 2,86 | 5,71 | Cruciani et al. (2007) | ||||
| Западная Азия | Юго-западная Турция | 40 | 2,5 | 2,5 | Cruciani et al. (2007) | |||||
| Западная Азия | Северо-восточная Турция | 41 | Cruciani et al. (2007) | |||||||
| Западная Азия | Юго-Восточная Турция | 24 | 4,17 | 4,17 | Cruciani et al. (2007) | |||||
| Западная Азия | Эрзурум Турецкий | 25 | 4 | 4 | Cruciani et al. (2007) | |||||
| Западная Азия | Центральная Анатолия | 61 | 6,56 | 1,64 | 4,92 | Cruciani et al. (2007) | ||||
| Западная Азия | Турецкие киприоты | 46 | 13,04 | 10,87 | 2,17 | Cruciani et al. (2007) | ||||
| Западная Азия | Сефардские турецкие | 19 | Cruciani et al. (2007) | |||||||
| Западная Азия | Палестинцы | 29 | 10,34 | 3,45 | 6,9 | Cruciani et al. (2007) | ||||
| Западная Азия | Друзы арабы | 28 | 10,71 | 10,71 | Cruciani et al. (2007) | |||||
| Западная Азия | Бедуины | 28 | 3,57 | 3,57 | Cruciani et al. (2007) | |||||
| Западная Азия | Сирийцы | 100 | 2 | 2 | Cruciani et al. (2007) | |||||
| Западная Азия | Курды из Ирака | 20 | Cruciani et al. (2007) | |||||||
| Западная Азия | Арабы из Арабских Эмиратов | 40 | 2,5 | 2,5 | Cruciani et al. (2007) | |||||
| Западная Азия | Оманит | 106 | 0,94 | 0,94 | Cruciani et al. (2007) | |||||
| Западная Азия | Адыгеи | 18 | Cruciani et al. (2007) | |||||||
| Западная Азия | Азери | 97 | 2,06 | 2,06 | Cruciani et al. (2007) |
Здесь основные субклады M78 по состоянию на июнь 2015 года. Внутри субклада E-M78, Trombetta et al. В 2015 году большая часть бывших хромосом E-M78 * была распределена по трем новым разным ветвям: E-V1083 *, E-V1477 и E-V259. Первый - сестра парагруппы кладов E-V22 и E-V13. Мутация V1477 определяет новую базальную ветвь, наблюдаемую только в одном образце из Северной Африки. Наконец, родственная родословная E-V12, определенная V264, включает E-V65 и новую центральноафриканскую линию, определенную V259. Редкая подгаплогруппа M78 E1b1b1a1-PF2186 наиболее часто встречается среди населения Тубу, населяющего Чад (21%).
Этот субклад E-M78, по-видимому, первым отделился от других (возник около 13,7-15,2 тыс. Лет назад). Согласно Cruciani et al. (2007), подлиния E-V12, вероятно, произошла от Северной Африки.
Недифференцированной линии передачи E-V12 * (не E-V32 или E-M224., Поэтому названный "E-V12 *") пик по частотам среди южных египтян (до 74,5%). Субклады также широко распространены в малых количествах в Северной Африке, так и в Европе, но с очень небольшими признаками в Азии, кроме Турции. Эти линии E-V12 * ранее были включены (вместе со многими линиями E-V22 *) в исходный (2004) "дельта-кластер" Cruciani et al., Который он определил с использованием профилей Y-STR. С открытием СНП Cruciani et al. (2007) сообщили, что самые высокие рекомендации V12 * были обнаружены в Египте, особенно в Южном Египте. Хассан и др. (2008) сообщают о значительном присутствии E-V12 * в соседнем Судане, включая 5/33 коптов и 5/39 нубийцев. E-V12 * составлял примерно 20% суданского E-M78. Они предполагают, что подклассы E-V12 и E-V22 E-M78 могут быть доставлены в Судан из их места происхождения в Северной Африке после прогрессирующего опустынивания Сахары около 6000–8000 лет назад. Внезапное изменение климата вынудить несколько неолитических культур / людей мигрировать на север в Средиземное море и на юг в Сахель и долину Нила. E-V12 * парагруппа также наблюдается в Европе (например, среди французских басков ) и Восточной Анатолии (например, Erzurum Турки ).
Небазальная подгаплогруппа E1b1b-V12 / E3b1a1 обнаружена с наибольшей популярностью среди различных популяций Восточной Африки, включая Гарре (74,1%), Габра (58,6%), Вата (55,6%), Борана. (50,0%), Sanye (41,7%), Beja (33,3%) и Rendille (29,0%).
E-M224 был обнаружен в Израиле среди йеменского населения (5%) и, по-видимому, является второстепенным субкладом.
Об его открытии было объявлено в Underhill et al. др. (2001) ошибка harvcoltxt: нет цели: CITEREFUnderhill_et_al.2001 (help ) и Шен и др. (2004) ошибка harvcoltxt: нет цели: CITEREFShen_et_al.2004 ( help ) найдено 1 из 20 йемени израильтян, которые они проверили. Cruciani et al. (2006) назвали M224 «редким и довольно малоинформативным »и не нашли xemplars.
 Распространение V-32 в Африке
Распространение V-32 в Африке Cruciani et al. (2007) предполагают, что этот субклад E-V12 возник в Северной Африке, а затем распространился дальше на юг до Африканского Рога, где он сейчас распространен. До открытия V32 Cruciani et al. (2004) ошибка harvcoltxt: нет цели: CITEREFCruciani_et_al.2004 (help ) указано на те же линии передачи, что и «гамма-кластер», который, по оценкам, возник около 8 500 лет назад. Они заявили, что "самые высокие частоты в трех кушитских говорящих: Борана из Кении (71,4%), оромо из Эфиопии (32,0%) и Сомали (52,2%). За пределами Восточной Африки он был обнаружен только у двух субъектов из Египта (3,6%) и у одного араба из Марокко ". Санчес и др. (2005) ошибка harvcoltxt: нет цели: CITEREFSanchez_et_al.2005 (help ) обнаружил, что это очень заметно у сомалийских мужчин и заявлено, что «это мужское сомалийское население» население Африканского Рога, связанное со к оромо в Эфиопии и Северной Кении (Боранас), «и что их гамма-кластерные линии» вероятно, были введены в сомалийскую популяцию 4000–5000 лет назад ». Совсем недавно Tillmar et al. (2009) ошибка harvcoltxt: нет цели: CITEREFTillmar_et_al.2009 (help ) типизировал 147 мужчин из Сомали по 12 локусам Y-STR и обнаружил, что 77% (113/147) имели типичные E- Гаплотипы V32. В настоящее время это самая высокая частота E-V32, обнаруженная в любой отдельной выборке. Аналогичным образом Hassan et al. (2008) в своем исследовании отметили, что это самая распространенная из подклассов E-M78, обнаруженных в Судане, особенно среди беджа, масалит. и Мех. Бежа, подобно сомалийцам и оромосам, говорят на афро-азиатском языке и живут вдоль «коридора» от Африканского Рога до Египта. Хассан и др. (2008) интерпретируют это как усиление «сильной корреляции между языками и генетическим разнообразием» и признаки родства между беджа и народами Африканского Рога, такими как амхара и оромо. С другой стороны, масалит и фур живут в Дарфуре и говорят на нило-сахарском языке. Авторы заметили в своем исследовании, «Масалит обладает наиболее высокими качествами гаплогруппы E-M78 и E-V32», что, по их мнению, предполагает «либо недавнее узкое место в популяции, либо близость к происхождению гаплогруппы». Однако совсем недавно Tillmar et al. (2009) ошибка harvcoltxt: нет цели: CITEREFTillmar_et_al.2009 (help ) набрал 147 мужчин из Сомали для 12 локусов Y-STR и обнаружил, что 77% (113 / 147) имели типичные гаплотипы E-V32. Это самая высокая частота E-V32, обнаруженная в любой отдельной выборке.
Данные STR из Cruciani et al. (2007) относительно E-V12 можно резюмировать следующим образом.
| Гаплотип | описание | YCAIIa | YCAIIb | DYS413a | DYS413b | DYS19 | DYS391 | DYS393 | DYS439 | A812S812S>DYS812S460S | ||
| E-V12 * | модальный | 19 | 22 | 22 | 22 | 13 | 10 | 13 | 11 | 11 | 9 | 13 |
| мин | 18 | 21 | 20 | 21 | 11 | 10 | 12 | 11 | 8 | 8 | 11 | |
| макс | 19 | 22 | 22 | 23 | 15 | 12 | 14 | 13 | 12 | 10 | 14 | |
| номер | 40 | 40 | 40 | 40 | 40 | 40 | 40 | 40 | 40 | 40 | 40 | |
| E-V32 | модальный | 19 | 21 | 22 | 23 | 11 | 10 | 13 | 12 | 10 | 10 | 13 |
| мин | 19 | 19 | 20 | 21 | 11 | 9 | 12 | 11 | 9 | 10 | 11 | |
| макс | 20 | 22 | 22 | 24 | 11 | 11 | 13 | 13 | 12 | 11 | 14 | |
| номер | 35 | 35 | 35 | 35 | 35 | 35 | 35 | 35 | 35 | 35 | 35 | |
| Все E-V12 | модальный | 19 | 22 | 22 | 23 | 11 | 10 | 13 | 11 | 11 | 10 | 13 |
| мин | 18 | 19 | 20 | 21 | 11 | 9 | 12 | 11 | 8 | 8 | 11 | |
| макс | 20 | 22 | 22 | 24 | 15 | 12 | 14 | 13 | 12 | 11 | 14 | |
| число | 75 | 75 | 75 | 75 | 75 | 75 | 75 | 75 | 75 | 75 | 75 | |
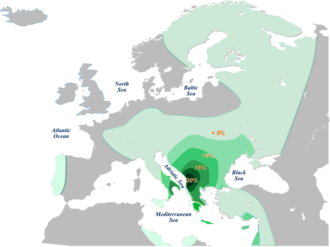 Распределение E-V13 согласно набору данных Cruciani (2007) и др., перечисленных выше
Распределение E-V13 согласно набору данных Cruciani (2007) и др., перечисленных выше Клада E-V13 эквивалентна «альфа-кластеру» E-M78, описанному в Cruciani и другие. (2004) ошибка harvcoltxt: нет цели: CITEREFCruciani_et_al.2004 (help ), и впервые была определена SNP V13 в Cruciani et al. (2006). Другой SNP известен для этой клады, V36, описанный в Cruciani et al. (2007). Все известные положительные тесты на V13 также положительны на V36. Таким образом, E-V13 в настоящее время считается «филогенетически эквивалентным» E-V36.
Гаплогруппа E-V13 - единственная линия передачи, которая показывает самых высоких частот за пределами Африки. Фактически, он составляет около 85% европейского хромосом E-M78 с клинальным паттерном частотного распределения от южной части Балканского полуострова (19,6%) до регистрации Европы (2,5%). Та же гаплогруппа также присутствует в более низких частотах в Анатолии (3,8%), Ближнем Востоке (2,0%) и Кавказе (1,8%). В Африке гаплогруппа E-V13 редко встречается только в Северной Африке с низкой низкой (0,9%).
— Cruciani et al. (2007)Согласно некоторым авторам, E-V13, по-видимому, возник в Греции или на южных Балканах, и его присутствие в остальной части Средиземного моря, вероятно, является следствием Греческая колонизация. В Европе E-V13 особенно распространен на Балканах и в некоторых частях Италии. В различных исследованиях особенно высокие частоты наблюдались у косовских албанцев (45,6%), македонских албанцев (34,4%), албанцев (32,29% Cruciani et al. (2007)), а также в некоторых частях Греции (около 35%). В целом, высокие частоты также были обнаружены в других регионах Греции, а также среди болгар, румын, македонцев и сербов.
в пределах Италия, частоты обычно выше в Южной Италии, с особенно высокими результатами, иногда наблюдаемыми в определенных областях; например, в Санта-Нинфа и Пьяцца Армерина в Сицилии. Высокие частоты, по-видимому, существуют также в некоторых северных областях, например, около Венеции, Генуи и Римини, а также на острове Корсика и регион Прованс на юге Франции, а также встречается в разбросанных и небольших количествах у ливийских евреев и Египта, но, скорее всего, это результат миграции из Европы или Ближнего Востока.
Среди древних образцов Loosdrecht et al. (2018) нашли одну окаменелость, несущую E-M78, в Grotte des Pigeons около Тафоральта в восточном Марокко. Скелет был непосредственно датирован периодом между 15 100 и 13 900 калиброванными годами до настоящего времени.
Очевидное перемещение линий E-M78 с Ближнего Востока в Европу и их последующее быстрое расширение делает его субклад E-V13 особенно интересным предметом для размышлений о древних миграциях людей.
Был сделан вывод, что именно в северо-восточной Африке, а не в восточной Африке, хромосомы E-M78 начали распространяться в другие регионы. Наиболее вероятен сценарий, что E-V13 возник в Западной Азии. Гипотеза состоит в том, что носители E-M78, лишенные мутации V13, покинули Африку и что слияние произошло позже на Ближнем Востоке / в Анатолии. Данные показывают, что западноазиатские носители V13 расширились в Европе не ранее 5300 лет назад. TMRCA европейской V13 датируется 4700–4000 лет назад. Филогенетический анализ предполагает, что европейский v13 распространился по Европе с Балкан в "быстрой демографической экспансии".
До этого мутация SNP, V13, по-видимому, впервые возникла в Западной Азии около 10 тысяч лет назад, но не широко распространена там, например, она встречается в больших количествах (>10% мужского населения) у турок-киприотов и друзов арабских родословных. Друзы считаются генетически изолированным сообществом и поэтому представляют особый интерес. Сигнатура ДНК STR некоторых из мужчин E-V13 среди них была первоначально отнесена к дельта-кластеру в Cruciani et al. (2004) ошибка harvcoltxt: нет цели: CITEREFCruciani_et_al.2004 (help ). Это означает, что Druze E-V13 сгруппировался вместе с большинством E-V12 и E-V22, а не с европейским E-V13, который в основном находился в альфа-кластере.
| гаплотип | описание | YCAIIa | YCAIIb | DYS413a | DYS413b | DYS19 | DYS391 | DYS393 | DYS439 | DYS460S <812812>DYS460S>A10 | ||
| Все E-V13 | модальные | 19 | 21 | 23 | 24 | 13 | 10 | 13 | 12 | 9 | 10 | 13 |
| Druze V13 | 1 | 19 | 21 | 23 | 23 | 13 | 10 | 13 | 13 | 11 | 9 | 12 |
| Druze V13 | 2 | 19 | 21 | 23 | 23 | 13 | 10 | 13 | 13 | 11 | 9 | 13 |
| Все E-V22 | модальные | 19 | 22 | 22 | 23 | 14 | 10 | 13 | 12 | 11 | 10 | 12 |
| Все E-V12 * | модальные | 19 | 22 | 22 | 22 | 13 | 10 | 13 | 11 | 11 | 9 | 13 |
Распространение и разнообразие V13 часто считается представлением сельскохозяйственных технологий во время неолита экспансии в Европу через Балканы. Гаплогруппа J2b (J-M12) также часто обсуждалась в связи с V13, как гаплогруппа с, казалось бы, очень похожим распределением и предысторией. (Нет единого мнения относительно обстоятельств или сроков его развития.)
Cruciani et al. (2007) говорит, что для этой географической области предполагалось как минимум четыре основных демографических события:
Последние две кажутся в пределах временного промежутка, возможного для V13 с учетом предполагаемого образования STR на среднеме. Воспользуемся связью с земледелием, человеческие останки, раскопанные в испанской погребальной пещере, датируемые примерно 7000 лет назад, оказались принадлежащими к этой гаплогруппе.
Однако более раннее проникновение в Европу также возможно. Battaglia et al. (2008), например, предполагают, что линия преемственности E-M78 *, являющаяся предком всех современных мужчин E-V13, быстро покинула родину в Южном Египте в более влажных условиях раннего голоцена ; прибыли на Балканы только с мезолитическими технологиями, а затем интегрировались с культурми неолита, которые позже прибыли на Балканы.
E-V13 в любом случае часто описывается в популяционной генетике как один из компонентов европейской генетической композиции, которая показывает относительно недавнюю связь популяций из Ближний Восток, входящий в Европу и предположительно связанный с внедрением новых технологий. из Африки в Евразию, которое было описано как «сигнал для отдельного позднего - Плейстоцен миграция из Африки. в Европу через Синай... которая не распределется в мДНК гаплогруппе.
После своего первоначального проникновения в Европу, произошло расселение с Балкан на остальную Европу. Также для этого движения существует широкий диапазон возможностей. Battaglia et al. (2008) предполагают, что субклад E-V13 E-M78 возник на месте в Европе, и предполагают, что первое крупное распространение E-V13 с Балкан с культурой неолита импрессированной посуды, которую часто называют Импрессой или Кардиал, могло происходить в направлении Адриатического моря.
Напротив, Кручиани и др. (2007 г.) подтверждает это предположение.) предполагают, что движение за пределы Балкан могло быть более недавним, чем 5300 лет назад. Авторы предполагают, что по большей части современные E-V13 оставалось на Балканах до Балканского бронзового века. Они считают, что «гаплогруппы E-V13 и J-M12 рассеялись в основном по речным водным путям, соединяющим южные Балканы с северной и центральной Европой». Перичич и др. (2005) ошибка harvcoltxt: нет цели: CITEREFPeričic_et_al.2005 (help ) предлагает реки Вардар-Морава-Дунай в качестве возможного пути распространения неолита в Центральную Европу. Берд (2007) предлагает еще более недавнее расселение с Балкан, примерно во времена Римской империи.
Согласно Lacan et al. (2011) ошибка harvcoltxt: цель отсутствует: CITEREFLacanKayserRicautBrucato2011 (help ), скелеты эпохи неолита (возраст ~ 7000 лет), которые были раскопаны в пещере Авелланер в Каталонии, включая северо -восточную Испанию особь мужского пола, несущая гаплогруппу E1b1b. Эта окаменелость принадлежит субкладу E1b1b1a1b (V13) и обладает идентичными гаплотипами, обнаруженными у современных европейцев (пять албанцев, два француза Прованса, два корсиканца, два боснийца, один итальянец, один сицилиец и один грек). Присутствие этой гаплогруппы в неолитической Испании предполагает, что она занимается с сельскохозяйственным комплексом эпохи неолита. Древний фермер также имел кладу мтДНК U5, раннюю европейскую материнскую гаплогруппу. Его аутосомные STR-маркеры также были наиболее типичными для Европы. Кроме того, образец был гомозиготным по C / C по LP-13910-C / T стойкость лактазы SNP, что указывает на непереносимость лактозы.
И E-M78 и J-M12 также использовались в исследованиях присутствующих на поисках свидетельств остающегося греческого происхождения в Афганистане и Пакистане, восходящем ко временам Александра Великого.
Обширный анализ Y-разнообразия у греков и трех пакистанских популяций - бурушо, калашей и патан - которые утверждают, что они произошли от греческих солдат, позволил нам сравните линии Y в этих популяциях повторно оцените предполагаемое греческое происхождение. Это исследование в целом, кажется, исключает большой вклад Греции в состав населения Пакистана, подтверждено предыдущие наблюдения. Однако это дает доказательные доказательства в поддержку греческого происхождения небольших частей патанов, о чем доказывает сеть кладов E и низкие генетические генетические дистанции между этими двумя популяциями.
— Firasat et al. (2006) ошибка harvcoltxt: нет цели: CITEREFFirasatKhaliqMohyuddinAisha2006 (help )Однако это исследование тестировалось только для M78, а не V13, типичного типа M78 с Балкан. Вместо этого «афганские родословные соотносятся с жителями Ближнего Востока и иранцами, но не с населением с Балкан»
Значительные частоты встречаемости E-V13 также наблюдались в городах Уэльса, около Честера (древний Deva Victrix ) в Англии и Шотландия. Старый торговый город Абергеле на северном побережье Уэльса, в частности, показано, что 7 из 18 протестированных местных жителей находились в этом происхождении (приблизительно 40%), как указано в Weale et al. (2002) ошибка harvcoltxt: нет цели: CITEREFWeale_et_a l.2002 (справка ).
Некоторые ученые (например, Стивен Берд (2007) ошибка harvcoltxt: нет цели: CITEREFSteven_Bird2007 (help ) приписывают присутствие E-V13 в Великом От Британии, особенно в наиболее часто встречающихся районах, до римских поселений в течение 1-4 веков н.э. Римская армия, включая людей балканского происхождения, включая фракийцев, иллирийцы и даки. В частности, Стивен Берд предлагает связь с современным регионом, включающим Косово, южную Сербию, северную Македонию и крайнюю северо-западную Болгарию - регион, соответствующую Римская провинция Верхняя Мезия, которая была определена Peričic et al. (2005) harvcoltxt error: no target: CITEREFPeričic_et_al.2005 (help ) как укрытие самая высокая частота этого субклада в мире.
Также примечательно, что E-V13, похоже, отсутствует в современной центральной Англии, особенно в Уэст-Мидлендсе и Южном Мидле. ands. Берд (2007) отмечает, что коллективный генетический профиль английских Мидлендс аналогичен голландской провинции Фрисландия, которая не была колонизирована Римом, но был, как и Англия, субъектом англосаксонского урегулирования. Так называемая «дыра E3b» в Центральной Англии, по словам Стивена Берда, может отражать замену населения - романо-британских людей англосаксами. Thomas et al. al. (2006) ошибка harvcoltxt: нет цели: CITEREFThomas_et_al.2006 (help ) повышает вероятность «апартеида », доминирования элиты социальных структур в англосаксонской Англии. Берд (2007) соглашается: «Дыра E3b» предполагает, что либо (а) массовое перемещение... романо-британского населения в результате вторжения, либо (б) существенная генетическая замена романо-британской популяции. Британская Y-ДНК через модель доминирования элиты ("апартеида")... Независимо от механизма, регион Центральной Англии... с его отсутствием гаплотипов E3b является областью, имеющей наиболее "поразительное сходство в распределении Y". -хромосомы "Фрисландия".
Хотя у большинства индивидуумов E-V13 не обнаруживаются какие-либо известные нижестоящие мутации SNP, и поэтому они классифицируются как E-V13 *, существует несколько признанных подкладов, все из которых могут быть очень маленькими. Это один из двух случаев, когда Karafet et al. (2008) ошибка harvcoltxt: нет цели: CITEREFKarafet_et_al.2008 (help ) заметил, что на момент написания этой статьи не было уверенности в том, что эти две клады действительно разделены («позиции этих мутации не были разрешены из-за отсутствия образца ДНК, содержащего производное состояние в V27 ").
Эта клада включает большинство кладовых, классифицированных в «дельта-кластере» Cruciani et al. (2004) ошибка harvcoltxt: нет цели: CITEREFCruciani_et_al.2004 (help ). Cruciani et al. (2006) позже отметили, что «хромосомы E-V22 и E-V12 * перемешаны и четко не дифференцированы по своим микросателлитным гаплотипам».
Этот субклад E-M78 «относительно распространен» в Африканском Роге и Египте с более высокой микросателлитной дисперсией (0,35 против 0,46, соответственно) в Египет. В статье, сообщающей об этой первой информации, Cruciani et al. (2007) описали это как необычное в Западной Азии и предложили Северо-Восточную Африку (Ливия / Египет) в качестве вероятного места происхождения этого субклада.
Самая высокая частота E-V22 до сих пор наблюдалась у кушитского -говорящего сахо населения центральной Эритреи со скоростью 88%. Известно, что сахо организованы в строгие патрилинейные и патрилокальные кланы. Была высказана гипотеза, что такого рода социальная структура может объяснять паттерны изменчивости, характеризующиеся низким разнообразием Y-хромосомы внутри групп и большим различием между группами.
Hassan et al. (2008) также сообщили о значительном присутствии в соседнем Судане, составляющем около 30% разнообразного диапазона линий передачи E-M78 страны в их исследовании, включая 8 из 26 фулани (около 31 %), широко рассредоточенный пастырский народ. E-V22 также присутствовал с гораздо меньшей частотой среди шиллуков (2 из 15 выборок, 13%) и динка (3 из 26, 11,5%) нилотов Южного Судана. Hassan et al. предполагают, что E-V22, как и E-V12, мог попасть в Судан из Северной Африки «после прогрессирующего опустынивания Сахары около 6000–8000 лет назад». Они добавляют, что поток генов в Судан «не только недавний (голоцен и далее), но и в значительной степени носит очаговый характер», и что «большинство носителей нило-сахарских языков, основные языковые семьи, на которых говорят в стране, демонстрируют очень мало признаков потока генов и демонстрируют низкий уровень миграции, за исключением нубийцев, которые, по-видимому, перенесли значительный поток генов из Азии и Европа вместе с Бежа. "
Другие частоты, указанные Cruciani et al. (2007) включает астурийцев (4,44% из 90 человек), сицилийцев (4,58% из 153 человек), марокканских арабов (7,27%, 55 человек), марокканские евреи (8%, 50 человек), Стамбул турки (5,71% из 35 человек) и палестинцы (6,9% из 29 человек). Cadenas et al. (2007) ошибка harvcoltxt: нет цели: CITEREFCadenas_et_al.2007 (help ) обнаружил 6,7% присутствия в ОАЭ.
Есть две признанные подклассы, которые, по-видимому, разделены, хотя Karafet (2008) harvcoltxt error: no target: CITEREFKarafet2008 (help ) отметил, что на момент написания этой статьи, «положения этих мутаций не были определены из-за отсутствия образца ДНК, содержащего производное состояние [...] V19».
Этот субклад, эквивалентный ранее классифицированному «бета-кластеру», находится на высоких уровнях в Магрибе. регионы далекого севера Африки. Cruciani et al. (2007) сообщают об уровне около 20% среди линий ливийских арабов и около 30% среди марокканских арабов. Он, кажется, менее распространен среди берберов, но все же присутствует на уровнях>10%. Авторы предполагают североафриканское происхождение этой линии. В Европе всего несколько особей были найдены в Италии и Греции. Результаты статьи можно резюмировать следующим образом...
| E-V65 | YCAIIa | YCAIIb | DYS413a | DYS413b | DYS19 | DYS391 | DYS393 | DYS439 | DYS460 | DYS461 | A10 |
| модальный | 19 | 21 | 21 | 23 | 13 | 10 | 13 | 10 | 10 | 11 | 13 |
| мин | 19 | 20 | 20 | 22 | 11 | 10 | 13 | 10 | 9 | 9 | 12 |
| макс | 21 | 21 | 22 | 23 | 14 | 11 | 14 | 11 | 11 | 12 | 13 |
| номер | 38 | 38 | 38 | 38 | 38 | 38 | 38 | 38 | 38 | 38 | 38 |
Capelli et al. al. (2009) ошибка harvcoltxt: нет цели: CITEREFCapelli_et_al.2009 (help ) изучил бета-кластер в Европе. Они обнаружили небольшие количества в Южной Италии, а также следы в Кантабрии, Португалии и Галисии, причем Кантабрия имела самый высокий уровень в Европе в их исследовании, 3,1% (5 из 161 человека).
Об открытии этого субклада было объявлено в Battaglia et al. (2008) Они обнаружили, что у 2 из 92 греков была эта мутация.
До 2002 года в академической литературе существовало по крайней мере семь систем именования для филогенетического дерева Y-хромосомы. Это привело к значительной путанице. В 2002 году основные исследовательские группы объединились и сформировали Консорциум Y-хромосомы (YCC). Они опубликовали совместный документ, в котором было создано единое новое дерево, которое все согласились использовать. Позже группа гражданских ученых, интересующихся популяционной генетикой и генетической генеалогией, сформировала рабочую группу для создания любительского дерева, стремясь быть, прежде всего, своевременным. В приведенной ниже таблице собраны все эти работы на основе знакового Дерева YCC 2002 года. Это позволяет исследователю, просматривающему ранее опубликованную литературу, быстро перемещаться между номенклатурами.
| YCC 2002/2008 (Сокращение) | (α) | (β) | (γ) | (δ) | (ε) | (ζ) | ( η) | YCC 2002 (от руки) | YCC 2005 (от руки) | YCC 2008 (от руки) | YCC 2010r (от руки) | ISOGG 2006 | ISOGG 2007 | ISOGG 2008 | ISOGG 2009 | ISOGG 2010 | ISOGG 2011 | ISOGG 2012 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| E-P29 | 21 | III | 3A | 13 | Eu3 | H2 | B | E* | E | E | E | E | E | E | E | E | E | E | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| E-M33 | 21 | III | 3A | 13 | Eu3 | H2 | B | E1 * | E1 | E1a | E1a | E1 | E1 | E1a | E1a | E1a | E1a | E1a | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 21 | III | 3A | 13 | Eu3 | H2 | B | E1a | E1a | E1a1 | E1a1 | E1a | E1a | E1a1 | E1a1 | E1a1 | E1a1 | E1a1 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| E-M75 | 21 | III | 3A | 13 | Eu3 | H2 | B | E2a | E2 | E2 | E2 | E2 | E2 | E2 | E2 | E2 | E2 | E2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 21 | III | 3A | 13 | Eu3 | H2 | B | E2b | E2b | E2b | E2b1 | - | - | - | - | - | - | - | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| E-P2 | 25 | III | 4 | 14 | Eu3 | H2 | B | E3 * | E3 | E1b | E1b1 | E3 | E3 | E1b1 | E1b1 | E1b1 | E1b1 | E1b1 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| E-M2 | 8 | III | 5 | 15 | Eu2 | H2 | B | E3a* | E3a | E1b1 | E1b1a | E3a | E3a | E1b1a | E1b1a | E1b1a | E1b1a1 | E1b1a1 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 8 | III | 5 | 15 | Eu2 | H2 | B | E3a1 | E3a1 | E1b1a1 | E1b1a1 | E3a1 | E3a1 | E1b1a1 | E1b1a1 | E1b1a1 | E1b1a1a1a | E1b1a1a1a | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 8 | III | 5 | 15 | Eu2 | H2 | B | E3a2<2993>a <335 | E1b1a2 | E1b1a2 | E3a2 | E3a2 | E1b1a2 | E1b1a2 | E1ba12 | удалено | удалено | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 8 | III | 5 | 15 | Eu2 | H2 | B | E3a3 | E3a3 | E1b1a3 | E1b1a3 | E3a3 | E3a3 | E1b1a3 | E1b1a3 | E1b1a3 | E1b1a1a1c | E1b1a1a1c | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 8 | 621>Eu2 | H2 | B | E3a4 | E3a4 | E1b1a4 | E1b1a4 | E3a4 | E3a4 | E1b1a4 | E1b1a4 | E1b1a4 | E1b1a1a1g1c | E1b1a1a1g1c | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 8 | III | 5 | 15 | Eu2 | H2 | B | E3a5 | E3a 335>E1b1a5 | E1b1a5 | E3a5 | E3a5 | E1b1a5 | E1b1a5 | E1b1a5 | E1b1a1a1d | E1b1a1a1d | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 8 | III | 5 | 15 | Eu2 | H2 | B | E3a6 | E3a6 | E1b1a6 | E1b1a6 | E3a6 | E3a6 | E1b1a6 | E1b1a6 | E1b1a6 | E1b1a1a1e | <3351a E1b1a 735>E-M3525 | III | 4 | 14 | Eu4 | H2 | B | E3b * | E3b | E1b1b1 | E1b1b1 | E3b1 | E3b1 | E1b1b1 | E1b1b1 | E1b1b1 | удалено | удалено | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| E-M78 | 25 | III | 4 | 14 | Eu4 | H2 | B | E3b1 * | E3b1 | E1b1b1a | E1b1b1a1 | E3b1a | E3b1a | E1b1b1a 299> | E1b1b1a | E1b1b1a | E1b1b1a1 | E1b1b1a1 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 25 | III | 4 | 14 | Eu4 | H2 | B | E3b1a | E3b1a 335>E1b1b1a3a | E1b1b1a1c1 | E3b1a3a | E3b1a3a | E1b1b1a3a | E1b1b1a3a | E1b1b1a3b | E1b1b1a3a <2992>E1b1b1a3a3a>E1b1b1a1c1E1b1b1a1c1 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 25 | III | 4 | 14 | Eu4 | H2 | B | E3b2* | E3b2 | E1b1b1b | E1b1b1b1b | E1b1b1b1b3 | E3b1b | E1b1b1b | E1b1b1b | E1b1b1b | E1b1b1b1 | E1b1b1b1a | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 25 | III4 | H2 | B | E3b2a | E3b2a | E1b1b1b1 | E1b1b1b1a | E3b1b1 | E3b1b1 | E1b1b1b1 | E1b1b1b1 | E1b1b1b1 | E1b1b1b1a | a <3351>E1b1b1b1aa <3351>III 623>Eu4H2 | B | E3b2b | E3b2b | E1b1b1b2 | E1b1b1b1 | E3b1b2 | E3b1b2 <2991a <3351>E3b1b2 <2991a <3351>299> | E1b1b1b2a | E1b1b1b2a | E1b1b1b2a | E1b1b1b1a2a | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| E-M123 | 25 | III | 4 | 14 | Eu4 <619 * <299b>E3 335>E3b3 | E1b1b1c | E1b1b1c | E3b1c | E3b1c | E1b1b1c | E1b1b1c | >E1b1b1cE1b1b1c | E1b1b1b2a | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 25 | III | 4 | 14 | Eu4 | H2 | B | E3b3a* | E3b3a | E1b1b1c1<2991> | E1b1b1c1<2991> | E3b1c1 | E3b1c1 | E1b1b1c1 | E1b1b1c1 | E1b1b1c1 | E1b1b1c1351>E2991>577>III | 4 | 14 | Eu4 | H2 | B | E3ba1 | E3b3a1 | E1b1b1c1a | E1b1b1c1a1 | E3b1c1a | E3b991c1a 335>E1b1b1c1a1 | E1b1b1c1a1 | E1b1b1c1a1 | E1b1b1c1a1 | E1b1b1b2a1a1 <74816>Исследования <20in>в следующих публикациях При создании дерева YCC были представлены g исследовательских групп в соответствии с их публикациями.
См. также
Genetics
субклады Y-ДНК E
Дерево остова Y-ДНК
ПримечанияСсылкиВнешние ссылки
Последняя правка сделана 2021-05-22 13:09:03
Содержание доступно по лицензии CC BY-SA 3.0 (если не указано иное). | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||