
Войти
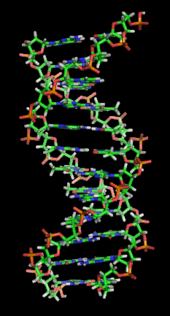
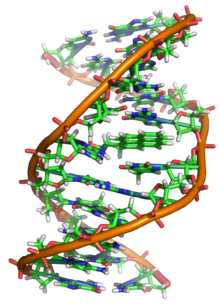
 Структура ДНК двойная спираль. Атомы в структуре имеют цветовую кодировку элемента, а подробные структуры двух пар оснований показаны в правом нижнем углу.
Структура ДНК двойная спираль. Атомы в структуре имеют цветовую кодировку элемента, а подробные структуры двух пар оснований показаны в правом нижнем углу.  Структура часть ДНК двойная спирали
Структура часть ДНК двойная спирали Дезоксирибонуклеиновая кислота ((![]() слушайте ); ДНК ) представляет собой молекула, состоящую из двух полинуклеотидных цепей, которые наматываются друг на друга с образованием двойной спирали, несущей генетические инструкции для функционирования, роста и воспроизводства всех известных организмов и многих вирусов. ДНК и рибонуклеиновая кислота (РНК) заменить собой нуклеиновые кислоты. Наряду с белками, липидами и сложными углеводами (полисахаридами ) нуклеиновые кислоты являются одним из четырех основных типов макромолекул, которые необходимы для всех известных формы жизни.
слушайте ); ДНК ) представляет собой молекула, состоящую из двух полинуклеотидных цепей, которые наматываются друг на друга с образованием двойной спирали, несущей генетические инструкции для функционирования, роста и воспроизводства всех известных организмов и многих вирусов. ДНК и рибонуклеиновая кислота (РНК) заменить собой нуклеиновые кислоты. Наряду с белками, липидами и сложными углеводами (полисахаридами ) нуклеиновые кислоты являются одним из четырех основных типов макромолекул, которые необходимы для всех известных формы жизни.
Две цепи ДНК известны как полинуклеотиды, поскольку они состоят из более простых мономерных единиц, называемых нуклеотидами. Каждый нуклеотид состоит из одного из четырех азотсодержащих азотистых оснований (цитозин [C], гуанин [G], аденин [A] или тимин [T]), сахар, называемый дезоксирибозой, и фосфатная группа. Нуклеотиды соединены друг с другом в цепи ковалентными связями (как фосфодиэфирная известная связь) между сахаром одного нуклеотида и фосфатом следующего, что приводит к чередующемуся сахару- фосфатный остов. Азотистые основания двух отдельных полинуклеотидных цепей связаны вместе в соответствии с правилами спаривания оснований (A с T и C с G) с водородными связями для образования двухцепочечной ДНК. Дополнительные азотистые основания разделены на две группы: пиримидины и пурины. В ДНК пиримидины - это тимин и цитозин; пурины - это аденин и гуанин.
Обе нити двухцепочечной ДНК хранят одинаковую биологическую информацию. Эта информация реплицируется, когда две нити разделяются. Большая часть ДНК (более 98% для человека) некодирует, что означает, что эти участки не образуют посадку для белковых последовательностей. Две цепи ДНК находятся в противоположных направлениях друг к другу и таким образом, антипараллельны. К каждому сахару присоединен один из четырех типов азотистых оснований (неформально, оснований). Именно последовательность из этих четырех азотистых оснований вдоль остова кодирует генетическую информацию. РНК цепи с использованием цепей ДНК в качестве матрицы в процессе, называемого транскрипцией, где основания ДНК заменяются основами, за исключением случая тимина (T), для которого РНК заменяет основания урацил (U). В рамках генетического кода эти цепи РНК определяют последовательность аминокислот в белках в процессе, называемом трансляцией.
. Внутри эукариотических клеток ДНК организована в длинные структуры, называемые хромосомы. Перед типичным делением клетки эти хромосомы дублируются в процессе репликации ДНК, полный набор хромосом для каждой дочерней клетки. Эукариотические организмы (животные, растения, грибы и простейшие ) хранят большую часть своей ДНК внутри ядро клетки как ядерная ДНК, а некоторые в митохондриях как митохондриальная ДНК или в хлоропластах как хлоропласт ДНК. Напротив, прокариоты (бактерии и археи ) хранят свою ДНК только в цитоплазме, в кольцевых хромосомах. Внутри эукариотических хромосом белки хроматина, такие как гистоны, уплотняют и организуют ДНК. Эти уплотняющие структуры направляют взаимодействие между ДНК и другими белками, помогая контролировать, какие части ДНК транскрибируются.
ДНК была впервые выделена Фридрихом Мишером в 1869 году. Ее молекулярная структура была идентифицирована Фрэнсисом Криком и Джеймсом Уотсоном на Лаборатория Кавендиша в Кембриджском университете в 1953 г., чьи усилия по построению моделей были основаны на данных дифракции рентгеновских лучей, полученные Раймондом Гослингом, который был аспирантом Розалинды Франклин в Королевском колледже Лондона. ДНК используется исследователями как молекулярный инструмент для изучения физических и теорий, таких как эргодическая теорема и теория эластичности. Уникальные свойства материала ДНК сделали ее привлекательную молекулы для материалов нанедов и инженеров, интересующихся микро- и производством. Среди заметных достижений в области - ДНК-оригами и гибридные материалы на этой основе ДНК.
 Химическая структура ДНК; водородные связи показаны пунктирными линиями
Химическая структура ДНК; водородные связи показаны пунктирными линиями ДНК полим представляет собой длинный ер, состоящий из повторяющихся звеньев, называемых нуклеотидами, каждая из которых обычно обозначается одной буквой: либо A, T, C, либо G. Структура ДНК динамична по всей длине, она способна скручиваться в плотные петли и другие формы. У всех видов он состоит из двух спиральных цепей, связанных друг с другом водородными связями. Обе цепи намотаны вокруг одной оси и имеют одинаковый шаг 34 грубрем (Å) (3,4 нанометров ). Пара цепочек имеет радиус 10 бритрем (1,0 нанометр). Согласно другому исследованию, при измерении другом в растворе длина цепи ДНК составляет от 22 до 26 нанометров (от 2,2 до 2,6 нанометров), длина одной нуклеотидной единицы - 3,3 Å (0,33 нм). Хотя каждый отдельный нуклеотид очень мал, полимер ДНК может быть очень большим и содержать миллионов, например, в хромосоме 1. Хромосома 1 является самой большой хромосомой человека с 220 миллионами пар оснований, и ее длина будет 85 мм, если ее выпрямить.
ДНК обычно не существует в виде одной нити, но вместо этого пара прядей, которые крепко скреплены. Эти две длинные нити наматываются друг на друга в форме двойной спирали . Нуклеотид содержит как сегмент основной цепи молекулы (который удерживает цепь вместе), так и азотистое основание (которое взаимодействует с другой цепью ДНК в спирали). Основание, связанное с сахаром, называется нуклеозидом, а основание, связанное с сахаром и одним или использованием фосфатными группами, называется нуклеотидом. Биополимер, состоящий из множества связанных нуклеотидов (как в ДНК), называется полинуклеотидом.
. Каркас цепи ДНК состоит из чередующихся фосфата и сахара группы. Сахар в ДНК - это 2-дезоксирибоза, которая представляет собой пентозный (пяти- углерод ) сахар. Сахара соединены вместе фосфатными группами, которые образуют фосфодиэфирные связи между третьим и пятым атомами углерода соседних сахарных колец. Они известны как 3'-конец (три основных конца) и 5'-конец (пять простых концов) атомы углерода, причем главный символ используется для отличия этих атомов углерода от те из основания, с которым дезоксирибоза образует гликозидную связь. Следовательно, любая цепь ДНК имеет один конец, на котором есть фосфатная группа, присоединенная к 5'-углеродному рибозы (5'-фосфорил), и другой конец, на котором есть свободная гидроксильная группа, присоединенная к 3'-углеродному атому рибозы. рибоза (3 'гидроксил). Ориентация углерода 3 'и 5' вдоль сахарно-фосфатного остова придает направленность (иногда называемую полярностью) каждой цепи ДНК. В двойной спирали нуклеиновой кислоты нуклеотидов в другой цепи противоположно их направление в цепи: цепи антипараллельны. Говорят, что асимметричные концы нитей ДНК имеют направленность пяти основных концов (5 ') и трех основных концов (3'), причем 5 'конец имеет концевую фосфатную группу, а 3' конец - концевую гидроксильную группу. Одно из основных различий между ДНК и РНК заключается в сахаре, при этом 2-дезоксирибоза в ДНК заменяется альтернативным пентозным сахаром рибозой в РНК.
 Участок ДНК. Основания лежат горизонтально между двумя спиралевидными цепями (анимированная версия ).
Участок ДНК. Основания лежат горизонтально между двумя спиралевидными цепями (анимированная версия ).Двойная спираль ДНК стабилизируется в основном двумя силами: водородными связями между нуклеотидами и взаимодействми стэкинга оснований среди ароматических азотистых оснований. Четыре основания, обнаруженные в ДНК: аденин (A), цитозин (C), гуанин (G) и тимин (T). Эти четыре основания присоединены к сахарному фосфату с образованием полного нуклеотида, как показано для аденозинмонофосфата. Аденин соединяется с тимином, пары гуанина с цитозином, образуя AT и GC пары оснований.
Азотистые основания подразделяются на два типа: пурины, A и G, которые представляют собой пяти- и шестичленные гетероциклические соединения и пиримидины, шестичленные кольца C и T. Пятое пиримидиновое азотистое основание, урацил (U), обычно занимае тим ина в РНК и от тимина отсутствием включения метильной группы в свое кольцо. Помимо РНК и ДНК, многие искусственные аналоги нуклеиновых кислот были созданы для использования в биотехнологии.
Модифицированные основания встречаются в ДНК. Первым из них был обнаружен 5-метилцитозин, который был обнаружен в геноме Mycobacterium tuberculosis в 1925 году. Причина присутствия этих неканонических оснований бактериальных вирусов (бактериофаги ), исключены рестрикционных ферментов, присутствующих в бактериях. Эта ферментная система действует, по крайней мере, частично как молекулярная иммунная система, защищающая бактерии от заражения вирусами. Модификации основ цитозина и аденина, наиболее распространенные и модифицированные ДНК, играют жизненно важную роль в эпигенетах контроле экспрессии генов у растений и животных.
Известно, что в ДНК встречается ряд неканонических оснований. Большинство из них являются модификациями канонических основ плюс урацил.
 ДНК и мелкие бороздки. Последний является сайтом связывания красителя Hoechst красителя 33258.
ДНК и мелкие бороздки. Последний является сайтом связывания красителя Hoechst красителя 33258. Двойные спиральные нити образуют основную цепь ДНК. Другая двойная спираль может быть обнаружена в промежутках или канавках между прядями. Эти пустоты возбуждают с парами оснований и соседствовать сайт связывания . Пряди не установлены симметрично друг другу, канавки имеют неодинаковый размер. Одна канавка, большая канавка, имеет ширину 22 имеетрем (Å), а другая, малая канавка, имеет ширину 12 Å. Ширина большой канавки означает, что края более доступны в большой канавке, чем в малой канавке. В результате белки, такие как факторы транскрипции , которые могут связываться со специфическими последовательностями в двухцепочечной ДНК, обычно контактируют со стороны оснований, выставленных в большой бороздке. Эта ситуация меняется в зависимости от необычной конформаций внутри клетки (см. Ниже), но большие и второстепенные бороздки всегда называются так, чтобы отражать различия в размерах, которые можно было бы увидеть, если ДНК скручивается обратно в обычную B-форму.
Двойная спирали ДНК каждая тип азотистых оснований на одной цепи связывается только с одним типом азотистых основ на другой цепи. Это называется комплементарной парой оснований. Пурины образуют водородные связи с пиримидинами, причем аденин связывается только с тимином по двум водородным связям, а цитозин связывается только с гуанином по трем водородным связям. Такое расположение двух нуклеотидов, связывающихся вместе по двойной спирали, называется парой оснований Уотсона-Крика. ДНК с высоким уровнем содержания GC более стабильна, чем ДНК с уровнем содержания GC. Пара оснований Хугстина - редкий вариант спаривания оснований. Водородные связи не установлены ковалентными, они могут быть разорваны и соединены относительно легко. Таким образом, две ДНК ДНК в двойной спирали могут быть разорваны, как застежка-молния, либо механической силой, либо высокой температурой. В результате комплементарности пар оснований информация в двухцепочечной системе спирали ДНК дублируется на каждую цепь, что жизненно важно для репликации ДНК. Это обратное специфическое взаимодействие между комплементарными парами основами важно для всех функций ДНК в критическом организме.
 |
 |
Как было указано выше, самая последняя молекула ДНК на самом деле представляет собой две полимерные нити, связанные друг с другом по спирали нековалентными связями; эта двухцепочечная (дцДНК ) структура в основном за счет соединений при укладке оснований внутри цепей, которые являются наиболее сильными для стеков G, C. Две могут разделиться - процесс, известный как плавление, - с образованием двух молекул одноцепочечной ДНК (оцДНК ). Плавление происходит при высокой температуре, низком уровне pH (низкий pH также плавит ДНК, но поскольку ДНК нестабильна из-за кислотной депуринизации, низкий pH используется редко).
Стабильность формы дцДНК зависит не только от содержания GC (% пар оснований G, C), но также от сохраняется (поскольку укладка зависит от последовательной), а также длину (более длинные молекулы более стабильны). Стабильность измерить разными способами; распространенным способом является «температура плавления», которая представляет собой температуру, при которой 50% молекул ds превращаются в молекулы ss; Температура плавления зависит от ионной силы и концентрации ДНК. В результате процента пар оснований GC, так и общая длина двойной спирали ДНК определяет силу ассоциации между двумя цепями ДНК. Имеют короткие спирали, содержащие GC. В биологии части двойной спирали ДНК, которые необходимо легко разделять, такие как TATAAT бокс Прибноу в некоторых промоторах, как правило, имеют высокое содержание AT, что облегчает выделение цепей.
В лаборатории значение этого взаимодействия можно измерить, найдя температуру, для разрыва половины водородных связей, температуру плавления (также называемую T m). Когда все пары оснований в двойной спирали ДНК плавятся, цепи разделяются и существуют в растворе как две полностью независимые молекулы. Эти одноцепочечные молекулы ДНК не единой общей формы, но некоторые конформации более стабильны, чем другие.
A ДНК-последовательность называется «смысловой» последовательностью, если она одинакова как копия информации РНК, которая транслируется в белок. Последовательность на противоположной цепи называется «антисмысловой» последовательностью. Как смысловые, так и антисмысловые последовательности могут существовать в разных частях одной и той же цепи (т.е. обе цепи могут содержать как смысловые, так и антисмысловые последовательности). Как у прокариот, так и у эукариот продуцируются антисмысловые продукты, поставляемые РНК, но функции этих РНК не совсем ясны. Одно из предположений состоит в том, что антисмысловые РНК участвуют в регулировании экспрессии генов посредством спаривания оснований РНК-РНК.
Несколько последовательностей ДНК в прокариотах и эукариотах и многое другое в плазмидах и вирусы стирают различие между смысловой и антисмысловой цепями из-за наличия перекрывающихся генов. В этих последовательностях происходит двойная функция, кодирующий один белок при считывании вдоль одной цепи и в противоположном направлении вдоль другой цепи. У бактерий это перекрытие может быть вовлечено в регуляцию транскрипции генов, тогда как вирусы перекрываются увеличенным количеством информации, которая может быть закодирована в небольшом вирусном геноме.
ДНК может быть скручена как веревка в процессе, называемая сверхспирализацией ДНК. Когда ДНК находится в «расслабленном» состоянии, нить обычно окружает ось двойной спирали каждые 10,4 пары оснований, но если ДНК скручена, нити становятся более плотными или более свободными. Если ДНК скручена в направлении спирали, это положительная суперспирализация, и основания удерживаются вместе более плотно. Если они скручены в противоположном направлении, это отрицательная сверхспирализация, и основания легче распадаются. В природе большая часть ДНК имеет небольшую отрицательную суперспирализацию, которая вводится ферментами, называемыми топоизомеразами. Эти ферменты также необходимы для снятия скручивающих напряжений во время таких процессов, как транскрипция и репликация ДНК.
 Слева направо, структуры ДНК A, B и Z
Слева направо, структуры ДНК A, B и Z ДНК существует во многих конформациях, которые включают в себя A-ДНК, B-ДНК и Z-ДНК, хотя непосредственно у функциональных механизмовов наблюдались только B-ДНК и Z-ДНК. Конформация, принимающая ДНК, от уровня гидратации, присутствующие ДНК, количества и направления суперспирализации, химических модификаций оснований, типа и концентрации концентраций металлов и концентрации полиаминов вре.
В первых опубликованных отчетах об A-ДНК рентгенограммах - а также B-ДНК - использовались анализы, основанные на преобразованиях Паттерсона, которые предоставил лишь ограниченный объем структурной информации для ориентированных волокон ДНК. Затем Уилкинс и др. В 1953 году предложили альтернативный анализ для in vivo B-ДНК рентгеновских дифракционных диаграмм рассеяния высокогидратированных ДНК в терминах квадратов функций Бесселя.. В том же журнале Джеймс Уотсон и Фрэнсис Крик представили свой анализ молекулярного моделирования дифрактограмм ДНК, чтобы предположить, что структура представляет собой двойную структуру. спираль.
Хотя форма B-ДНК наиболее распространена в условиях, встречающихся в клетках, это не четко определенная конформация, а семейство родственных конформаций ДНК, которые имеют высокий уровень гидратации, присутствующих в клетках. Соответствующие картины дифракции рентгеновских лучей и рассеяния характерны для молекулярных паракристаллов со степенью степени беспорядка.
По сравнению с B-ДНК форма A-ДНК более широкая правая спираль с неглубокой широкой малой бороздкой и более узкой и глубокой большой бороздкой. Форма A встречается в нефизиологических условиях в частично дегидратированных образцах ДНК, тогда как в клетке она может образовываться в гибридных парах ДНК и цепей РНК, а также в комплексах фермент-ДНК. Сегменты ДНК, основания которых были химически модифицированы посредством метилирования, могут претерпевать большее изменение конформации и принимать Z-. Здесь нити вращаются вокруг оси спирали по левой спирали, противоположной более распространенной формы B. Эти необычные структуры могут участвовать в регуляции транскрипции, связывающими Z-ДНК. Исследование 2020 года пришло к выводу, что ДНК повернулась вправо из-за ионизации космическими лучами.
В течение многих лет экзобиологи предполагали существование теневая биосфера, постулируемая микробная биосфера Земли, использует совершенно другие биохимические и молекулярные процессы, чем в известном в настоящее время жизни. Одним из предложений было существование форм жизни, которые используют мышьяк вместо фосфора в ДНК. В 2010 году было объявлено о возможности наличия бактерии GFAJ-1, хотя результаты были оспорены, и данные о том, что бактерия активно предотвращает включение мышьяка в основную цепь ДНК. и другие биомолекулы.
На концах линейных хромосом находятся специализированные области ДНК, называемые теломерами. Основная функция этих возможностей - использование ферментов клеток, которые реплицируют концы хромосом с использованием теломераза, поскольку ферменты, которые обычно реплицируют ДНК, не могут копировать крайние 3'-концы хромосом. Эти специализированные хромосомные заглушки также обеспечивают защиту концы ДНК и не позволяют системам репарации ДНК в клетке рассматривать как повреждения, которые необходимо исправить. В клетках человека теломеры обычно предоставляют собой отрезки одноцепочечной ДНК, содержит несколько тысяч повторов простой TTAGGG.
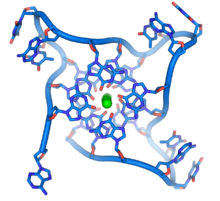 Квадруплекс ДНК, образованный теломерными повторами. Петлевидная конформация остова ДНК сильно отличается от типичной спирали ДНК. Зеленые сферы в центре предложения ионы калия.
Квадруплекс ДНК, образованный теломерными повторами. Петлевидная конформация остова ДНК сильно отличается от типичной спирали ДНК. Зеленые сферы в центре предложения ионы калия. Эти богатые гуаниные структуры могут стабилизировать концы хромосом, образуя структуры из четырех основных единиц, а не обычные пары оснований, обнаруженные в других молекулах ДНК. Здесь четыре гуаниновых основания, известные как тетрада гуанина, образуют плоскую пластину. Эти плоские блоки с четырьмя базами затем складываются на друга, образуя друг друга стабильную G-квадруплекса. Эти структуры стабилизируются за счет водородных связей между краями оснований и хелатирования иона металла в центре каждой четырехосновной единицы. Также могут быть сформированы другие структуры, при этом центральный набор из четырех основ происходит либо из одной нити, сложной вокруг основы, либо из нескольких различных параллельных нитей, каждая из которых вносит одну основу в центральную структуру.
В дополнение к этим сложным друг структурам теломеры также образуют большие петлевые структуры, называемые петлями теломер или Т-петлями. Здесь одноцепочечная ДНК закручивается в длинный круг, стабилизированный белками, связывающими теломеры. В самом конце Т-петли одноцепочечная теломерная ДНК удерживается на участке двухцепочечной ДНК цепью теломер, разрушая двухспиральную ДНК и спаривание оснований с одной из двух цепей. Эта трехцепочечная структура называется петлей с ущерба или D-петлей.
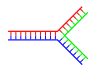 |  |
| Одна ветвь | Несколько ветвей |
В ДНК изнашивание происходит, когда некомплементарные области существуют на конце другой комплементарной двойной цепи ДНК. Разветвленная ДНК может возникнуть, если вводится третья цепь ДНК, и она содержит новые области, способные гибридизоваться с потрепанными областями ранее существовавшей двойной цепи. Хотя простейший пример разветвленной ДНК включает только три цепи ДНК, также возможны комплексы, включающие дополнительные цепи и несколько ответвлений. Разветвленная девушка может Роман в нанотехнологии для создания геометрических фигур, см. Раздел использование в технологии ниже.
Было синтезировано несколько искусственных азотистых основ, которые успешно включены в восьмиосновный аналог ДНК, названный Hachimoji DNA. Названные S, B, P и Z, эти искусственные основания способны взаимодействовать друг с другом с другом образом (S - B и P - Z), поддерживать двойную спирали ДНК и транскрибироваться в РНК. Их подразумевает, что в четырех естественных азотистых основаниях, развившихся на Земле, нет ничего особенного.
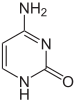 |  |  |
| цитозин | 5-метилцитозин | тимин |
На экспрессию генов влияет то, как ДНК упакована в хромосомах, обрабатываемую, называемую хроматин. Модификации оснований могут быть вовлечены в упаковку с участками, которые имеют низкую экспрессию генов или не ее, обычно содержат высокие уровни метилирования оснований цитозина. Обработка ДНК и ее влияние на экспрессию генов также может происходить путем ковалентных модификаций ядра белка гистона, вокруг которого ДНК обернута в структуре хроматина, или путем ремоделирования, осуществляемого комплексами ремоделирования хроматина (см. Ремоделирование хроматина ). Кроме того, существует перекрестная помеха метилированием ДНК и модификацией гистонов, поэтому они могут координированно влиять на хроматин и экспрессию генов.
Например, метилирование цитозина дает 5-метилцитозин, что важно для X-инактивации хромосом. Средний уровень метилирования оценки между организмом: у червя Caenorhabditis elegans отсутствует метилирование цитозина, в то время как у позвоночных уровень метилирования выше, причем до 1% их ДНК содержит 5-метилцитозин. Несмотря на важность 5-метилцитозина, он может дезаминировать, оставляя тиминовое основание, поэтому метилированные цитозины, особенно склонны к мутациям. Другие модификации оснований включают метилирование аденина у бактерий, присутствие 5-гидроксиметилцитозина в мозге и гликозилирование урацила с образованием «J-основания» в кинетопластидах.
 A ковалентного аддукта между метаболически активированной формой бензо [a] пирена, основной мутаген в табачные дыме, ДНК
A ковалентного аддукта между метаболически активированной формой бензо [a] пирена, основной мутаген в табачные дыме, ДНК ДНК могут быть использованы различными видами мутагенов, которые изменяют последовательность ДНК. Мутагены включают окислители, алкилирующие агенты, а также высокоэнергетическое электромагнитное излучение, такое как ультрафиолет свет и рентгеновское излучение. Тип повреждения ДНК зависит от типа мутагена. Например, УФ-свет может повредить ДНК, образуя димеры тимина, которые представляют собой поперечные связи между пиримидиновыми основаниями. С другой стороны, окислители, такие как свободные радикалы или перекись водорода, вызывают различные повреждения, модификации формы оснований, особенно гуанозина, и двухцепочечные разрывы. Типичная человеческая клетка содержит около 150 000 оснований, поврежденных окислением. Из этих двух окислительных повреждений наиболее опасными являются поддающиеся восстановлению точечные мутации, вставки, делеции в ДНК. последовательность и хромосомные транслокации. Эти мутации могут вызвать рак. Из-за ограничений, присущих механизмам восстановления ДНК, если бы люди жили достаточно долго, у всех в итоге разовьется рак. Повреждения ДНК, возникают в природе, из-за нормальных клеточных процессов, которые производят активные формы кислорода, гидролитической активности клеточной воды и т.д., также часто встречаются. Хотя большая часть этих повреждений восстанавливается, в любой клетке остаются некоторые повреждения ДНК. Эти повреждения ДНК накапливаются с возрастом в постмитотических тканях млекопитающих. Это по-видимому, накопление основной причиной старения.
Многие мутагены помещаются в пространство между двумя соседними парами основаниями, это называется интеркаляцией. Большинство интеркаляторов - это ароматические и плоские молекулы; примеры включают бромид этидия, акридины, дауномицин и доксорубицин. Чтобы интеркалятор поместился между парами оснований, основания разделяются, искаженная цепь ДНК за счетручивания двойной спирали. Это подавляет как транскрипцию, так и репликацию ДНК, вызывая токсичность и мутации. В результате интеркаляторы ДНК могут быть канцерогенами, а в случае талидомида - тератогеном. Другие, такие как эпоксид бензо [a] пирендиола и афлатоксин, образуют аддукты ДНК, которые вызывают ошибки в репликации. Тем не менее, из-за их способности ингибировать транскрипцию и репликацию ДНК, другие аналогичные токсины также используются в химиотерапии для подавления быстрорастущих раковых клеток.
 Расположение эукариот ядерной ДНК в хромосомах
Расположение эукариот ядерной ДНК в хромосомах ДНК обычно встречается в виде линейных хромосом у эукариот и кольцевых хромосом в прокариоты. Набор хромосом в клетке составляет ее геном ; геном человека содержит 3 миллиарда пар оснований ДНК, ДНК в 46 хромосомах. Информация, переносимая ДНК, содержащая в последовательность фрагментов ДНК, называемых генами. Передача генетической информации в генах достигается посредством комплементарного спаривания оснований. Например, при транскрипции, когда клетка использует информацию в гене, последовательность ДНК копируется в комплементарную последовательность РНК за счет притяжения между ДНК и правильными нуклеотидами РНК. Обычно эта копия РНК затем используется для создания соответствующей последовательности белка в процессе, называемом трансляцией, который зависит от того же взаимодействия между нуклеотидами РНК. В качестве альтернативы клетка может просто копировать свою генетическую информацию в процессе, называемом репликацией ДНК. Подробности этих функций описаны в других статьях; здесь основное внимание уделяется взаимодействиям между ДНК и другими молекулами, которые обеспечивают функцию генома.
Геномная ДНК плотно и упорядоченно упакована в процессе, называемом конденсацией ДНК, чтобы соответствовать небольшим доступным объемам клетки. У эукариот ДНК находится в ядре клетки, с небольшими количествами в митохондриях и хлоропластах. У прокариот ДНК удерживается внутри тела неправильной формы в цитоплазме, называемого нуклеоидом. Генетическая информация в геноме хранится в генах, и полный набор этой информации в организме называется его генотипом. Ген - это часть ДНК, которая влияет на определенную характеристику организма. Гены содержат открытую рамку считывания, которая может быть транскрибирована, и регуляторные последовательности, такие как промоторы и энхансеры, которые контролируют транскрипцию открытых рамка чтения.
У многих видов только небольшая часть общей последовательности генома кодирует белок. Например, только около 1,5% генома человека состоит из кодирующих белок экзонов, при этом более 50% ДНК человека состоит из некодирующих повторяющихся последовательностей. Причины наличия такого большого количества некодирующей ДНК в геномах эукариот и необычайные различия в размере генома или значении C среди видов представляют собой долгую загадка, известная как "загадка значения C ". Однако некоторые последовательности ДНК, которые не кодируют белок, могут по-прежнему кодировать функциональные некодирующие молекулы РНК, которые участвуют в регуляции экспрессии генов.
 РНК-полимеразы T7 (синий) вырабатывает мРНК (зеленый) из матрицы ДНК (оранжевый)
РНК-полимеразы T7 (синий) вырабатывает мРНК (зеленый) из матрицы ДНК (оранжевый) Некоторые некодирующие ДНК не структурные роль в хромосомах. Теломеры и центры обычно содержат несколько генов, но они важны для функций и стабильности хромосом. Распространенной формой некодирующей ДНК у людей являются псевдогены, которые заменены собой копии генов, которые были отключены в результате мутации. Эти наблюдения обычно показывают собой молекулярные окаменелости, хотя иногда они могут служить необработанным генетическим материалом для создания новых генов в процессе дупликации генов и расхождение.
Ген - это последовательность ДНК, которая содержит генетическую информацию и может влиять на фенотип организма. Внутри гена последовательность оснований вдоль цепи ДНК определяет последовательность информационной РНК, которая затем определяет одну или несколько белковых последовательностей. Взаимосвязь между нуклеотидными последовательностями генов и аминокислотными последовательностями белков определяется правилами трансляции, которые вместе известны как генетический код. Генетический код состоит из трехбуквенных «слов», называемых кодонами, образованных из последовательности из трех нуклеотидов (например, ACT, CAG, TTT).
При транскрипции кодоны гена копируются в информационную РНК с помощью РНК-полимеразы. Эта копия РНК затем декодируется рибосомой , которая считывает последовательность РНК путем спаривания оснований информационной РНК с транспортной РНК, которая несет аминокислоты. Поскольку имеется 4 основания в 3-буквенных комбинациях, существует 64 возможных кодона (4 комбинации). Они кодируют двадцать стандартных аминокислот , что дает большинству аминокислот более одного возможного кодона. Есть также три «стоповых» или «бессмысленных» кодона, обозначающих конец кодирующей области; это кодоны TAA, TGA и TAG.
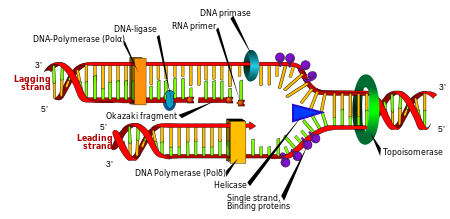 Репликация ДНК: Двойная спираль разматывается геликаза и топоизомераза. Затем одна ДНК-полимераза производит копию ведущей цепи. Другая ДНК-полимераза связывается с отстающей цепью. Этот фермент прерывистые сегменты (называемые фрагментами Окадзаки ) до того, как ДНК-лигаза соединяет их вместе.
Репликация ДНК: Двойная спираль разматывается геликаза и топоизомераза. Затем одна ДНК-полимераза производит копию ведущей цепи. Другая ДНК-полимераза связывается с отстающей цепью. Этот фермент прерывистые сегменты (называемые фрагментами Окадзаки ) до того, как ДНК-лигаза соединяет их вместе. Деление клетки необходимо для роста организма, но когда клетка делится, она должна реплицировать ДНК в своем геноме, чтобы две дочерние клетки имели ту же генетическую информацию, что и их родительские. Двухцепочечная структура ДНК обеспечивает простой механизм репликации ДНК. Здесь две цепи разделяются, и каждая последовательность комплементарной ДНК воссоздается с помощью средства , называемого ДНК-полимеразой. Этот фермент комплементарную цепь, находя правильное основание посредством комплементарного спаривания оснований и подключая его к исходной цепи. ДНК-полимеразы-полимеразы могут использовать ДНК в направлении от 5 'до 3', для копирования антипараллельных цепей двойная спирали используются различные механизмы. Таким образом, основание на старой цепи определяет, какое основание появляется на новой цепи, и в итоге получает идеальную копию своей ДНК.
Голая внеклеточная ДНК (еДНК), большая часть которой высвобождается в результате гибели клеток, почти повсеместно присутствует в окружающей среде. Его способность в почве может достичь 2 мкг / л, а в естественной водной среде может достичь 88 мкг / л. Для eDNA были предложены различные возможные функции: она может участвовать в горизонтальном переносе гена ; он может обеспечить питательными веществами; и он может действовать как буфер для набора или титрующего инструмента или антибиотиков. Внеклеточная ДНК работает как функциональный компонент внеклеточного матрикса в биопленках нескольких видов бактерий. Он может действовать как определение распознавания, регулировать распространение типов клеток в биопленке; это может образования биопленки; и это может быть прочной прочности биопленки и устойчивости к биологическому стрессу.
Внеклеточная ДНК плода обнаружена в крови матери и может быть секвенирована для определения большого количества информации о развивающемся плоде.
Под названием ДНК окружающей среды эДНК получила все большее распространение в качестве инструмента исследования экологии, мониторинга перемещений и присутствия видов в воде., воздух или на суше, и оценка биоразнообразия местности.
Все функции ДНК зависят от взаимодействия с белками. Эти белковые взаимодействия могут быть неспецифическими, или белок специфически связываться с одной последовательностью ДНК. Ферменты также могут связываться с ДНК, и из них особенно важны полимеразы, которые копируют последовательность оснований ДНК при транскрипции и репликации ДНК.
 Взаимодействие ДНК (оранжевый) с гистонами (синий). Аминокислоты этих связываются с кислыми фосфатными группами ДНК.
Взаимодействие ДНК (оранжевый) с гистонами (синий). Аминокислоты этих связываются с кислыми фосфатными группами ДНК. Структурные белки, связывающие ДНК, являются понятными примерами неспецифических взаимодействий ДНК-белок. Внутри хромосом ДНК находится в комплексах со структурными белками. Эти белки организуют ДНК в компактную структуру, называемую хроматином. У эукариот эта структура включает связывание ДНК с комплексом основных белков, называемых гистонами, тогда как у прокариот задействованы несколько типов белков. Гистоны образуют дискообразный комплекс, называемый нуклеосомой, который содержит два полных витка двухцепочечной ДНК, обернутой вокруг ее поверхности. Эти неспецифические вызовы образуются через основные остатки в гистонах, образуя ионные связи с кислым сахарно-фосфатным остальным ДНК и, таким образом, в образовании степени независимы от оснований. Химические модификации этих аминокислотных остатков включают метилирование, фосфорилирование и ацетилирование. Эти химические изменения изменяют силу взаимодействия ДНК и гистонами, ДНК более или менее доступной для факторов транскрипции и изменяя скорость транскрипции. Другие неспецифические ДНК-связывающие белки в хроматине включают белки группы с высокой подвижностью, которые связываются с изогнутой или искаженной ДНК. Эти белки важны для изгиба массивов нуклеосом и их упорядочения в более крупные структуры, составляющие хромосомы.
Отдельной группой ДНК-связывающих белков ДНК-связывающие белки, которые специфически связывают одноцепочечную ДНК. У людей репликационный белок А является наиболее изученным членом этого семейства и используется в процессе, в котором двойная спираль разделяется, включая репликацию, рекомбинацию и репарацию ДНК. Эти связывающие белки, по-видимому, стабилизируют однонитевую ДНК и защищают ее от образования петель или разрушения нуклеазами.
 Репрессор лямбда спираль-поворот-спираль транскрипция фактор, связанный с его ДНК-мишенью
Репрессор лямбда спираль-поворот-спираль транскрипция фактор, связанный с его ДНК-мишенью Напротив, белки эволюционировали, чтобы связываться с конкретными последовательностями ДНК. Наиболее интенсивно из них изучаются различные факторы транскрипции, которые обеспечивают собой белки, регулирующие транскрипцию. Каждый фактор транскрипции связывается с одним набором последовательностей ДНК и активирует или ингибирует транскрипцию генов, которые имеют эти последовательности, близкие к их промоторам. Факторы транскрипции делают это двумя способами. Во-первых, они могут связываться с РНК-полимеразой, ответственной за транскрипцию, либо напрямую, либо через другие белки-медиаторы; это определяет местонахождение полимеразы на промоторе и позволяет ей начать транскрипцию. Альтернативно, факторы транскрипции могут связывать ферменты, которые модифицируют гистоны на промоторе. Это изменяет доступность матрицы ДНК для полимеразы.
Эти ДНК-мишени могут встречаться по всему геному организма, изменения в активности типа фактора транскрипции влиять на тысячи генов. Следовательно, эти белки часто являются мишенями процессов передачи сигнала, которые контролируют ответы на изменения окружающей среды или клеточную дифференцировку и развитие. Специфичность взаимодействия этих факторов транскрипции ДНК обусловлена множественными контактами белков с краями оснований ДНК, что позволяет им «читать» последовательность ДНК. Большинство этих основий происходит в большой бороздке, где основания наиболее доступны.
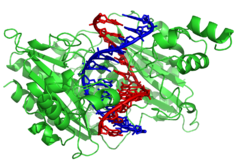 рестрикционный фермент EcoRV (зеленый) в комплексе с его субстратной ДНК
рестрикционный фермент EcoRV (зеленый) в комплексе с его субстратной ДНК Нуклеазы - это ферменты, которые разрезают цепи ДНК, катализируя гидролиз сложного фосфодиэфира. облигации. Нуклеазы, гидролизующие нуклеотиды в концах цепей ДНК, называются экзонуклеазами, а эндонуклеазами разрезаются внутри цепей. Наиболее часто используемые нуклеазы в молекулярной биологии - это рестрикции эндонуклеазы, которые разрезают ДНК по определенным последовательностям. Например, фермент EcoRV, показанный слева, распознает последовательность из 6 оснований 5'-GATATC-3 'и делает разрез на горизонтальной линии. В природе эти ферменты защищают бактерии от инфекции фагом, переваривая ДНКага, когда она проникает в бактериальную клетку, действуя как часть <системы539>рестрикционной модификации. В технологии специфичные для последовательности нуклеазы используются в молекулярном клонировании и ДНК-фингерпринте.
Ферменты, называемые ДНК-лигазами, могут соединяться с разрезанными или разорванными цепями ДНК. Лигазы особенно важны в репликации отстающей цепи ДНК, поскольку они объединяют короткие ДНК-матрицы, образующиеся на репликационной вилке, в полную копию ДНК-матрицы. Они также используются в репарации ДНК и генетической рекомбинации..
Топоизомеразы являются ферментами с нуклеазной и лигазной активностью. Эти белки изменяют количество сверхспирализации в ДНК. Некоторые из этих ферментов работают, разрезая спираль ДНК. фермент затем запечатывает разрыв ДНК. Другие типы данных ферментов представать одну спираль ДНК. Топоизомеразы необходимы для многих процессов с участием ДНК, таких как репликация и транскрипция ДНК.
Геликазы - это белки, которые предоставляют собой тип молекулярного мотора. Они используют химическую энергию в нуклеозидтрифосфатах, преимущественно в аденозинтрифосфате (АТФ), для разрыва водородных связей между основаниями и раскручивания двойной спирали ДНК на одиночные нити. Эти ферменты необходимы для обеспечения процессов, где ферментам необходим доступ к основанию ДНК.
Полимеразы ферментируют собой менты, которые синтезируют полинуклеотидные цепи из нуклеозидтрифатов. Последовательность их продуктов создается на основе алгоритма полинуклеотидных схем, которые называются шаблонами. Эти ферменты функционируют путем многократного добавления нуклеотида к 3 'гидроксильной группе на конце растущей полинуклеотидной цепи. Как следствие, все полимеразы в направлении от 5 'до 3'. В активном сайте этих ферментов входящие пары оснований нуклеозидтрифосфата в матрицу: это позволяет полимеразам точно синтезировать комплементарную цепь матрицы. Полимеразы классифицируются по типу используемого шаблона.
ДНК-зависимые ДНК-зависимые ДНК-полимеразы копии полинуклеотидных цепей ДНК. Для сохранения биологической информации важно, чтобы последовательность оснований в каждой копии системы была точно основана на матрице. Многие ДНК-полимеразы обладают активностью корректуры. Здесь полимераза распознает случайные ошибки синтеза из-за отсутствия спаривания оснований между несовпадающими нуклеотидами. При обнаружении несоответствия активируется активность экзонуклеазы от 3 'до 5' и удаляется неправильное основание. ДНК-зажимы-полимеразы, называемые реплисомой, которые содержат несколько дополнительных субъединиц, таких как ДНК-зажим или геликазы.
РНК-зависимые ДНК. -полимеразы. Предоставьте собой специализированный класс полимераз, копирующие последовательности цепи РНК в ДНК. Они включают обратную транскриптазу, которая представляет собой вирусный фермент, участвующий в инфицировании клеток ретровирусами, и теломеразу, которая требуется для репликации теломер. Например, обратная транскриптаза ВИЧ - это фермент для репликации вируса СПИДа. Теломераза - необычная полимераза, поскольку она содержит собственную матрицу РНК как часть своей структуры. Он синтезирует теломеры на концах хромосом. Теломеры предотвращают влияние концов соседних хромосом и защищают концы хромосом от повреждений.
Транскрипция осуществляется с помощью-зависимой РНК-полимеразы, которая копирует ДНК в РНК. Чтобы начать транскрибирование гена, РНК-полимераза связывается с последовательностью ДНК, называемой промотором, и разделяет цепи ДНК. Затем он копирует последовательность гена в транскрипт информационный РНК до тех пор, пока не достигнет области ДНК, называемой терминатором, где она останавливается и отделяется от ДНК. Как и в случае использования ДНК-зависимых ДНК-полимераз человека, РНК-полимераза II, фермент, транскрибирующий большинство генов в геноме человека, действует как часть большого белкового комплекса с множеством регуляторных и отдельные субъединицы.
 |
 |
 Рекомбинация включает разрыв и воссоединение двух хромосом (M и F) с образованием двух перестроенных хромосом (C1 и C2).
Рекомбинация включает разрыв и воссоединение двух хромосом (M и F) с образованием двух перестроенных хромосом (C1 и C2). Обычно спираль ДНК не взаимодействует с другими сегментами ДНК, а в клетках человека разные хромосомы даже занимают области в ядре, называемые «территориями хромосом ». Это физическое состояние разных хромосомных способностей ДНК функционирует как стабильное хранилище информации, поскольку происходит из немногих случаев хромосомного кроссовера , который происходит во время полового размножения, когда происходит генетическая рекомбинация. Хромосомный кроссовер - это когда две спирали ДНК разрываются, меняют местами участок и затем соединяются.
Рекомбинация позволяет хромосомам обмениваться генетической информацией и повторять новые комбинации генов, увеличивает естественного отбора и может иметь важное значение для быстрой эволюции новых белков. Генетическая рекомбинация также может участвовать в репарации ДНК, особенно в ответе клетки на двухцепочечные разрывы.
Наиболее распространенная схема хромосомного кроссовера является гомологичная рекомбинация, при которой две участвующие хромосомы имеют общие похожие последовательности. Негомологичная рекомбинация может приводить к повреждению клеток, поскольку она может вызывать хромосомные транслокации и генетические аномалии. Реакция рекомбинации катализируется ферментами, известными как рекомбиназы, например, RAD51. Первым этапом рекомбинации является двухцепочечный разрыв, вызванный либо эндонуклеазой, либо повреждением ДНК. Затем серия стадий частично катализируемых рекомбиназой приводит к соединению двух спиралей по крайней мере соединением Холлидея, в котором сегмент одной цепи в каждой спирали отжигается с комплементарной цепью в другой спираль. Соединение Холлидея - это тетраэдрическая структура соединения, которое можно перемещать по паре хромосом, меняя одну нить на другую. Затем реакция рекомбинации останавливают расщеплением соединения и повторным лигированием высвободившейся ДНК. Только нити одинаковой полярности обменивают ДНК во время рекомбинации. Существует два типа расщепления: расщепление восток-запад и расщепление север-юг. Расщепление север-юг разрезает обе нити ДНК, тогда как расщепление восток-запад сохраняет одну нить ДНК нетронутой. Образование соединения Холлидея во время рекомбинации делает возможным генетическое разнообразие, обмен генами на хромосомах и экспрессию вирусных геномов дикого типа.
ДНК содержит генетическую информацию, которая позволяет всем формам жизни функционировать, расти и воспроизводиться. Как неясно, как долго за 4 миллиарда лет истории жизни ДНК выполняла эту функцию, как было высказано предположение, что самые ранние формы жизни могли использовать РНК в качестве своего генетического материала. РНК могла выступать в качестве передаваемой информации, так и выполнять катализ как часть рибозимов. Этот древний мир РНК, где нуклеиновая кислота могла бы найти как для катализа, так и для генетики, возможно, повлияла на эволюцию текущего генетического кода, основанного на четырехотидальных основаниях. Это могло бы произойти, поскольку количество различных оснований в таком организме - это компромисс между небольшим количеством оснований, увеличивающим точность репликации, и большим количеством оснований, увеличивающим каталитическую эффективность рибозимов. Однако нет прямых доказательств существования древних генетических систем, поскольку восстановление ДНК из большинства окаменелостей невозможно, поскольку ДНК выживает в окружающей среде менее одного миллиона лет и медленно распадается на короткие фрагменты в растворе. Были сделаны заявления о более древней ДНК, в первую очередь сообщение о выделении жизнеспособной бактерии из кристалла соли 250 миллионов лет, но эти утверждения спорны.
Строительные блоки ДНК (аденин, гуанин и родственные органические молекулы ) могли быть образованы внеземными цивилизациями в космическом пространстве. Комплексная ДНК и РНК органические соединения из life, включая урацил, цитозин и тимин, также были сформированы в лаборатории в условиях, имитирующих условия, обнаруженные в космическом пространстве, с использованием исходных химикатов, таких как пиримидин, обнаруженных в метеоритах. Пиримидин, как и полициклические ароматические углеводороды (ПАУ), наиболее богатое углеродом химическое вещество, обнаруженное во вселенной, возможно, образовалось в красных гигантах или в межзвездных космическая пыль и газовые облака.
Были разработаны методы очистки ДНК от организмов, таких как фенол -экстракция хлороформа, и манипуляции с ней в лаборатории, такие как рестрикционные расщепления и полимеразная цепная реакция. В современной биологии и биохимии эти методы интенсивно используются в технологии рекомбинантных ДНК. Рекомбинантная ДНК представляет собой последовательность ДНК, созданную человеком, которая была собрана из других последовательностей ДНК. Их можно трансформировать в организмы в форме плазмид или в соответствующем формате, используя вирусный вектор. Полученные генетически модифицированные организмы могут использоваться для производства таких продуктов, как рекомбинантные белки, использоваться в медицинских исследованиях или выращиваться в сельском хозяйстве.
Судмедэксперты могут использовать ДНК в крови, сперме, коже, слюне или волосах найден на месте преступления, чтобы идентифицировать совпадающую ДНК человека, например преступника. Этот процесс формально называется профилирование ДНК, также называемое дактилоскопией ДНК. При профилировании ДНК длины различных участков повторяющейся ДНК, таких как короткие тандемные повторы и минисателлиты, сравниваются между людьми. Этот метод обычно является чрезвычайно надежным методом выявления совпадающей ДНК. Однако идентификация может быть затруднена, если место происшествия заражено ДНК нескольких человек. Анализ ДНК был разработан в 1984 году британским генетиком сэром Алеком Джеффрисом и впервые использован в судебной медицине для осуждения Колина Пичфорка в 1988 году убийства Эндерби.
Разработка криминалистики и возможность теперь получить генетическое соответствие мельчайших образцов крови, кожи, слюны или волос привели к пересмотру многих дел. Теперь могут быть обнаружены доказательства, которые были невозможны с научной точки зрения во время первоначального исследования. В сочетании с отменой закона о двойной угрозе в некоторых местах это может позволить возобновить дела, если предыдущие судебные процессы не смогли предоставить достаточных доказательств, чтобы убедить присяжных. Людей, обвиняемых в серьезных преступлениях, могут попросить предоставить образец ДНК для сопоставления. Самая очевидная защита от совпадений ДНК, полученных судебно-медицинской экспертизой, - это утверждение о перекрестном заражении улик. Это привело к тщательным и строгим процедурам рассмотрения новых дел о серьезных преступлениях.
ДНК-профилирование также успешно используется для точной идентификации жертв массовых несчастных случаев, тел или частей тел в серьезных авариях, а также отдельных жертв в массовых военных захоронениях путем сопоставления с членами семьи.
ДНК-профилирование также используется в ДНК-тестировании на отцовство, чтобы определить, является ли кто-либо биологическим родителем или бабушкой или дедушкой ребенка с вероятностью отцовства обычно 99,99%, когда предполагаемый родитель биологически связан ребенку. Обычные методы секвенирования ДНК применяются после рождения, но есть новые методы проверки отцовства, пока мать еще беременна.
дезоксирибозимы, также называемые ДНКзимы или каталитическая ДНК были впервые обнаружены в 1994 году. В основном они представляют собой одноцепочечные последовательности ДНК, выделенные из большого пула случайных последовательностей ДНК с помощью комбинаторного подхода, называемого in vitro отбором или систематической эволюцией лигандов с помощью экспоненциальное обогащение (SELEX). ДНКзимы катализируют множество химических реакций, включая расщепление РНК-ДНК, лигирование РНК-ДНК, фосфорилирование-дефосфорилирование аминокислот, образование углерод-углеродных связей и т.д. ДНКзимы могут увеличивать каталитическую скорость химических реакций до 100000000000 раз по сравнению с некаталитической реакцией. Наиболее широко изученный класс ДНКзимов - это типы, расщепляющие РНК, которые использовались для обнаружения различных ионов металлов и создания терапевтических агентов. Сообщалось о нескольких металлоспецифичных ДНКзимах, включая ДНКзим GR-5 (специфичный для свинца), ДНКзим CA1-3 (специфичный для меди), ДНКзим 39E (специфичный для уранила) и ДНКзим NaA43 (специфичный для натрия). ДНКзим NaA43, который, как сообщается, более чем в 10000 раз селективен к натрию по сравнению с ионами других металлов, был использован для создания сенсора натрия в клетках в реальном времени.
Биоинформатика включает в себя разработку методов для хранения, сбора данных, поиска и обработки биологических данных, включая данные последовательности нуклеиновой кислоты ДНК . Это привело к широко применяемым достижениям в информатике, особенно в алгоритмах поиска строк, машинном обучении и теории баз данных. Алгоритмы поиска или сопоставления строк, которые находят вхождение последовательности букв внутри более крупной последовательности букв, были разработаны для поиска конкретных последовательностей нуклеотидов. Последовательность ДНК может быть выровнена с другими последовательностями ДНК для идентификации гомологичных последовательностей и локализации конкретных мутаций, которые делают их отличными. Эти методы, особенно множественное выравнивание последовательностей, используются при изучении филогенетических взаимоотношений и функции белков. Наборы данных, представляющие ценность последовательностей ДНК целого генома, например, полученные в рамках Human Genome Project, трудно использовать без аннотаций, определяющих расположение генов и регуляторных элементов на каждой хромосоме. Области последовательности ДНК, которые имеют характерные паттерны, связанные с генами, кодирующими белки или РНК, могут быть идентифицированы с помощью алгоритмов поиска генов, которые позволяют исследователям прогнозировать наличие определенных генных продуктов и их возможные функции в организме еще до того, как они были выделены экспериментально. Также можно сравнивать целые геномы, что может пролить свет на эволюционную историю конкретного организма и позволить исследовать сложные эволюционные события.
 Структура ДНК слева (показана схематическая) будет самособираться в структуру, визуализированную атомно-силовой микроскопией справа. ДНК-нанотехнология - это область, которая стремится создавать наноразмерные структуры с использованием свойств молекулярного распознавания молекул ДНК. Изображение из Стронг, 2004 г..
Структура ДНК слева (показана схематическая) будет самособираться в структуру, визуализированную атомно-силовой микроскопией справа. ДНК-нанотехнология - это область, которая стремится создавать наноразмерные структуры с использованием свойств молекулярного распознавания молекул ДНК. Изображение из Стронг, 2004 г..ДНК-нанотехнология использует уникальные свойства молекулярного распознавания ДНК и других нуклеиновых кислот для создания самособирающихся разветвленных комплексов ДНК с полезными свойствами. Таким образом, ДНК используется как структурный материал, а не как носитель биологической информации. Это привело к созданию двумерных периодических решеток (как на основе плиток, так и с использованием метода ДНК-оригами ) и трехмерных структур в форме многогранников. Также были продемонстрированы наномеханические устройства и алгоритмическая самосборка, и эти структуры ДНК были использованы для моделирования расположения других молекул, таких как наночастицы золота и стрептавидин. белки.
Поскольку ДНК собирает мутации с течением времени, которые затем передаются по наследству, она содержит историческую информацию, и, сравнивая последовательности ДНК, генетики могут сделать вывод об эволюционной истории организмов, их филогения. Эта область филогенетики - мощный инструмент в эволюционной биологии. Если сравнивать последовательности ДНК внутри вида, популяционные генетики могут узнать историю конкретных популяций. Это может быть использовано в различных исследованиях: от экологической генетики до антропологии.
ДНК как устройство хранения информации имеет огромный потенциал, поскольку имеет гораздо более высокую плотность хранения по сравнению с электронными устройствами. Однако высокая стоимость, чрезвычайно медленное время чтения и записи (задержка памяти ) и недостаточная надежность препятствовали его практическому использованию.
 Джеймс Уотсон и Фрэнсис Крик (справа), соавторы модели двойной спирали, с Маклином Маккарти (слева)
Джеймс Уотсон и Фрэнсис Крик (справа), соавторы модели двойной спирали, с Маклином Маккарти (слева)  Карандашный рисунок двойной спирали ДНК, сделанный Фрэнсисом Криком в 1953 году
Карандашный рисунок двойной спирали ДНК, сделанный Фрэнсисом Криком в 1953 году ДНК был впервые выделен швейцарским врачом Фридрихом Мишером, который в 1869 году обнаружил микроскопическое вещество в гное выброшенных хирургических повязок. Поскольку он находился в ядрах клеток, он назвал его нуклеином. В 1878 г. Альбрехт Коссель выделил небелковый компонент «нуклеина», нуклеиновой кислоты, а затем выделил его пять основных азотистых оснований.
. В 1909 г. Фебус Левен идентифицировал основание, сахар и фосфатно-нуклеотидная единица РНК (затем называемая «дрожжевой нуклеиновой кислотой»). В 1929 году Левен идентифицировал сахар дезоксирибозы в «нуклеиновой кислоте тимуса» (ДНК). Левен предположил, что ДНК состоит из цепочки из четырех нуклеотидных единиц, связанных друг с другом через фосфатные группы («тетрануклеотидная гипотеза»). Левен думал, что цепь короткая, а основания повторяются в фиксированном порядке. В 1927 году Николай Кольцов предположил, что унаследованные признаки будут унаследованы через «гигантскую наследственную молекулу», состоящую из «двух зеркальных цепей, которые будут реплицироваться полуконсервативным образом, используя каждую цепочку в качестве шаблона». В 1928 году Фредерик Гриффит в своем эксперименте обнаружил, что черты «гладкой» формы пневмококка могут быть переданы «грубой» форме тех же бактерий. путем смешивания убитых «гладких» бактерий с живой «грубой» формой. Эта система дала первое четкое предположение о том, что ДНК несет генетическую информацию.
В 1933 году, изучая яйца девственных морских ежей, Жан Бреше предположил, что ДНК находится в ядре клетки и что РНК присутствует исключительно в цитоплазме. В то время считалось, что «нуклеиновая кислота дрожжей» (РНК) встречается только в растениях, а «нуклеиновая кислота тимуса» (ДНК) - только у животных. Последний считался тетрамером с функцией буферизации клеточного pH.
В 1937 году Уильям Эстбери создал первые дифракционные рентгенограммы, которые показали, что ДНК имеет правильную структуру..
В 1943 году Освальд Эйвери вместе с коллегами Колином МакЛаудом и Маклин Маккарти идентифицировали ДНК как трансформирующую принцип, подтверждающий предположение Гриффита (эксперимент Эйвери – Маклауда – Маккарти ). Роль ДНК была подтверждена в 1952 году, когда Альфред Херши и Марта Чейз в эксперименте Херши-Чейза показали, что ДНК является генетическим материалом фага энтеробактерий T2.
 A синяя бляшка снаружи The Eagle pub в память о Крике и Уотсоне
A синяя бляшка снаружи The Eagle pub в память о Крике и Уотсоне В конце 1951 года Фрэнсис Крик начал работать с Джеймсом Уотсоном в Кавендишской лаборатории в Кембриджском университете. В феврале 1953 года Линус Полинг и Роберт Кори предложили модель нуклеиновых кислот, содержащую три переплетенные цепи, с фосфатами около оси и основаниями снаружи. В мае 1952 года Раймонд Гослинг аспирант, работавший под руководством Розалинд Франклин, сделал снимок дифракции рентгеновских лучей, помеченный как «Фото 51. ", при высоком уровне гидратации ДНК. Эта фотография была предоставлена Уотсону и Крику Морисом Уилкинсом и имела решающее значение для получения ими правильной структуры ДНК. Франклин сказал Крику и Ватсону, что костяк должен быть снаружи. До этого у Линуса Полинга, Уотсона и Крика были ошибочные модели с цепями внутри и основанием, направленными наружу. Ее идентификация пространственной группы для кристаллов ДНК показала Крику, что две цепи ДНК были антипараллельными.
. В феврале 1953 года Уотсон и Крик завершили свою модель, которая теперь принята как первая правильная. модель двойной спирали ДНК. 28 февраля 1953 года Крик прервал обеденный перерыв для посетителей в The Eagle pub в Кембридже, чтобы объявить, что он и Уотсон « открыли секрет жизни».
25 апреля 1953 года в выпуске журнала Nature была опубликована серия из пяти статей, в которых приводилась структура ДНК Ватсона и Крика с двойной спиралью и подтверждающие ее доказательства. Об этой структуре сообщалось в письме, озаглавленном «МОЛЕКУЛЯРНАЯ СТРУКТУРА ЯДЕРНЫХ КИСЛОТ. Структура для нуклеиновой кислоты дезоксирибозы», в котором они сказали: «Это не ускользнуло от нашего внимания, что определенная пара, которую мы постулировали, сразу же предполагает возможный механизм копирования генетических материал ". Затем последовало письмо Франклина и Гослинга, которое было первой публикацией их собственных данных дифракции рентгеновских лучей и их оригинального метода анализа. Затем последовало письмо Уилкинса и двух его коллег, в котором содержался анализ структуры рентгеновских лучей B-ДНК in vivo и поддерживалось присутствие in vivo структуры Уотсона и Крика.
В 1962 г. после смерти Франклина Уотсон, Крик и Уилкинс совместно получили Нобелевскую премию по физиологии и медицине. Нобелевские премии присуждаются только живым лауреатам. Продолжаются споры о том, кому следует отдать должное за открытие.
В влиятельной презентации в 1957 году Крик изложил центральную догму молекулярной биологии, которая предсказала взаимосв язь между ДНК, РНК, и белки, и сформулировал «гипотезу адаптера». Окончательное подтверждение механизма репликации, подразумеваемого двойной спиральной структурой, за которой последовал в 1958 году эксперимент Мезельсона – Шталя. Дальнейшая работа Крика и его сотрудников показала, что генетический код основан на неперекрывающихся триплетах оснований, называемых кодонами, что позволяет Хар Гобинд Хорана, Роберт У. Холли и Маршалл Уоррен Ниренберг для расшифровки генетического кода. Эти результаты представляют собой рождение молекулярной биологии.
| В Викицитатнике есть цитаты, связанные с: ДНК |
| Викиверситет имеет обучающие ресурсы по ДНК |
| На Викискладе есть медиафайлы, связанные с ДНК. |
{kind=link}