
Войти
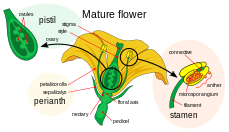 Части цветка
Части цветка  Двойное оплодотворение
Двойное оплодотворение  Двойное оплодотворение у Arabidopsis
Двойное оплодотворение у Arabidopsis Двойное оплодотворение представляет собой сложный механизм оплодотворения цветковых растений (покрытосеменных ). Этот процесс включает соединение женского гаметофита (мегагаметофита, также называемого зародышевым мешком ) с двумя мужскими гаметами (спермой). Он начинается, когда пыльцевое зерно прилипает к рыльцу плодолистика, женской репродуктивной структуры цветка. Затем пыльцевое зерно впитывает влагу и начинает прорастать, образуя пыльцевую трубку, которая проходит вниз к завязи сквозным слоем. Затем кончик пыльцевой трубки входит в яичник и проходит через отверстие микропиле в семяпочке. Пыльцевая трубка продолжает выпуск двух сперматозоидов мегагаметофита.
В неоплодотворенной яйцеклетке имеется 8 клеток, расположенных в виде 3 + 2 + 3 (сверху вниз), то есть 3 антиподальных клетки, 2 полярных центральных клетки, 2 синергид и 1 яйцеклетка. Один сперматозоид оплодотворяет яйцеклетку, а другой сперматозоид соединяется с двумя полярными ядрами большой центральной клетки мегагаметофита. Гаплоидный сперматозоид и гаплоидное яйцо объединяются, образуя диплоидную зиготу, процесс называется сингамией, в то время как другой сперматозоид и два гаплоидных полярных ядра большого центральная клетка мегагаметофита образует триплоидное ядро (тройное слияние ). Некоторые растения могут образовывать полиплоидные ядра. Затем большая клетка гаметофита разовьется в эндосперм, богатую питательными веществами ткань, обеспечивающую питание развивающегося эмбриона. Яичник, окружающий семяпочки, развивается в плод, который защищает семена и может функционировать для их распространения.
Два центральных материнских ядра клетки (полярные ядра), которые вносят вклад в эндосперм, возникают в результате митоза из тот же единственный мейотический продукт, из которого произошло яйцо. Материнский вклад в генетическую конституцию триплоидного эндосперма вдвое больше, чем у эмбриона.
В исследовании растения Arabidopsis thaliana, проведенного в 2008 году, миграция мужских ядер внутри женской гаметы в слиянии с женскими ядрами впервые была задокументирована с использованием in vivo визуализация. Также были определены некоторые гены, участвующие в процессе миграции и слияния.
Сообщалось о доказательствах двойного оплодотворения у Gnetales, которые не являются цветущими семенными растениями.
Двойное оплодотворение было открыто более века назад Сергеем Наващиным и Гриньяром в Киеве, Российской Империи, и Леон Гиньяр в Франции. Каждый из них сделал открытие независимо от другого. Lilium martagon и Fritillaria tenella были использованы в первых наблюдениях за двойным оплодотворением, которые были сделаны с использованием классической световой микроскоп. Из-за ограничений светового микроскопа оставалось много безответных вопросов относительно процесса двойного оплодотворения. Однако с развитием электронного микроскопа на многие вопросы были даны ответы. В частности, наблюдения, проведенные группой W. Jensen, показали, что мужские гаметы не имеют каких-либо клеточных стенок и что плазматическая мембрана гамет находится близко к плазматической мембране. клетки, которая их окружает внутри пыльцевого зерна.
Гораздо более рудиментарная форма двойного оплодотворения происходит при половом размножении отряда голосеменных, широко известных как Gnetales. В частности, это событие было задокументировано как в Ephedra, так и в Gnetum, подмножестве Gnetophytes. В Ephedra nevadensis одна двуядерная сперматозоид откладывается в яйцеклетку. После первоначального оплодотворения второе ядро сперматозоидов направляется на оплодотворение дополнительного ядра яйцеклетки, обнаруженного в цитоплазме яйцеклетки. У большинства других семенных растений это второе «ядро вентрального канала» обычно оказывается функционально бесполезным. У Gnetum gnemon в цитоплазме самки внутри женского гаметофита имеются многочисленные свободные ядра яйца. После проникновения зрелого женского гаметофита через пыльцевую трубку женская цитоплазма и свободные ядра перемещаются, чтобы окружить пыльцевую трубку. Из двухъядерных клеток сперматозоидов выделяются два ядра сперматозоидов, которые затем соединяются со свободными ядрами яйцеклетки с образованием двух жизнеспособных зигот, что является гомологичной характеристикой между семействами Ephedra и Gnetum. В обеих семьях второе оплодотворение дает дополнительный диплоидный эмбрион. Позднее этот лишний эмбрион выкидывается, что приводит к синтезу только одного зрелого эмбриона. Продукт дополнительного оплодотворения в эфедре не питает первичный эмбрион, поскольку женский гаметофит отвечает за обеспечение питательными веществами. Более примитивный процесс двойного оплодотворения у голосеменных приводит к образованию двух диплоидных ядер, заключенных в одной яйцеклетке. Это отличается от состояния покрытосеменных, которое приводит к разделению яйцеклетки и эндосперма. Сравнительное молекулярное исследование генома G. gnemon показало, что гнетофиты более тесно связаны с хвойными деревьями, чем с покрытосеменными. Отказ от гипотезы об антофитах, которая определяет, что gnetales и покрытосеменные являются сестринскими таксонами, приводит к предположению, что процесс двойного оплодотворения является продуктом конвергентной эволюции и возник независимо среди гнетофитов и покрытосеменных растений.
Двойное оплодотворение in vitro часто используется для изучения молекулярных взаимодействий, а также других аспектов слияния гамет у цветковых растений. Одним из основных препятствий на пути к двойному оплодотворению in vitro мужских и женских гамет является ограничение сперматозоидов в пыльцевой трубке и яйцеклетки в зародышевом мешке. Контролируемое слияние яйцеклетки и сперматозоидов уже было достигнуто с растениями мака. Прорастание пыльцы, проникновение пыльцевых трубок и процессы двойного оплодотворения протекали нормально. Фактически, этот метод уже использовался для получения семян различных цветковых растений и получил название «оплодотворение из пробирки».
женский гаметофит, мегагаметофит, который участвует в двойном оплодотворении покрытосеменных, который является гаплоидным, называется зародышевым мешком. Он развивается в семяпочке, окруженной яичником у основания плодолистика. Мегагаметофит окружен (одним или) двумя покровами, которые образуют отверстие, называемое микропиле. Megagametophyte, который, как правило, гаплоидный, берет свое начало от (обычно диплоидный ) мегаспоры материнская клетка, которая также называется megasporocyte. Следующая последовательность событий варьируется в зависимости от конкретного вида, но у большинства видов происходят следующие события. Мегаспороцит подвергается мейотическому делению клеток, образуя четыре гаплоидных мегаспоры. Выживает только одна из четырех образующихся мегаспор. Эта мегаспора проходит три раунда митотического деления, в результате чего образуются семь клеток с восемью гаплоидными ядрами (центральная клетка имеет два ядра, называемые полярными ядрами). Нижний конец зародышевого мешка состоит из гаплоидной яйцеклетки, расположенной в середине двух других гаплоидных клеток, называемых синергидами. Функции синергии в привлечении и направлении пыльцевой трубки к мегагаметофиту через микропиле. На верхнем конце мегагаметофита находятся три антиподальные клетки.
Мужские гаметофиты или микрогаметофиты, которые участвуют в двойном оплодотворении, содержатся в пыльцевых зернах. Они развиваются в микроспорангиях, или пыльцевых мешочках, пыльников на тычинках. Каждый микроспорангий содержит диплоидные микроспоры материнские клетки или микроспороциты. Каждый микроспороцит подвергается мейозу, образуя четыре гаплоидных микроспоры, каждая из которых в конечном итоге может развиться в пыльцевое зерно. Микроспора подвергается митозу и цитокинезу, чтобы произвести две отдельные клетки, генеративную клетку и трубочную клетку. Эти две клетки помимо стенки спор составляют незрелое пыльцевое зерно. По мере созревания мужского гаметофита генеративная клетка переходит в трубочную клетку, а генеративная клетка подвергается митозу, производя две сперматозоиды. После созревания пыльцевых зерен пыльники раскрываются, высвобождая пыльцу. Пыльца переносится ветром или животными опылителями к пестику другого цветка и откладывается на рыльце. По мере прорастания пыльцевого зерна трубчатая клетка производит пыльцевую трубку, которая удлиняется и простирается вниз по длинной части плодолистика до яичника, где его сперматозоиды высвобождаются в мегагаметофит. Отсюда происходит двойное оплодотворение.