
Войти
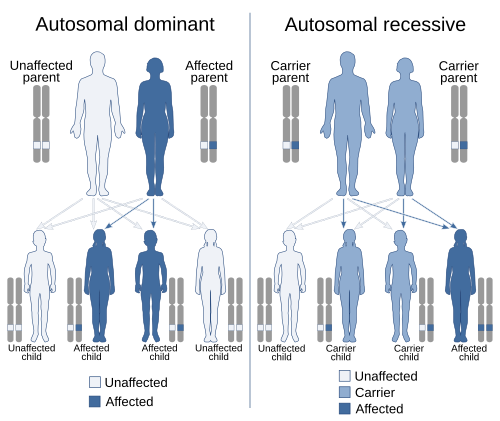 Аутосомно-доминантное и аутосомно-рецессивное наследование, два наиболее распространенных образца менделевского наследования. аутосома - это любая хромосома, отличная от половой хромосомы.
Аутосомно-доминантное и аутосомно-рецессивное наследование, два наиболее распространенных образца менделевского наследования. аутосома - это любая хромосома, отличная от половой хромосомы.В генетике, доминирование - это явление одного варианта (аллель ) гена на хромосоме маскирует или отменяет эффект другого варианта того же гена на другую копию хромосомы. Первый вариант называется доминантным, а второй рецессивным . Это состояние наличия двух разных вариантов одного и того же гена на каждой хромосоме изначально вызвано мутацией в одном из генов, либо новым (de novo), либо. Термины аутосомно-доминантный или аутосомно-рецессивный используются для описания вариантов генов на неполовых хромосомах (аутосомы ) и связанных с ними черт, в то время как на половые хромосомы (аллосомы) называются Х-сцепленным доминантным, Х-сцепленным рецессивным или Y-сцепленным ; у них есть шаблон наследования и представления, который зависит от пола как родителя, так и ребенка (см. Связь полов ). Поскольку существует только одна копия Y-хромосомы, Y-сцепленные признаки не могут быть ни доминантными, ни рецессивными. Кроме того, существуют другие формы доминирования, такие как неполное доминирование, при котором вариант гена имеет частичный эффект по сравнению с тем, когда он присутствует на обеих хромосомах, и совместное доминирование в какие разные варианты на каждой хромосоме проявляют связанные с ними черты.
Доминирование не присуще аллелю или его признакам (фенотип ). Это строго относительный эффект между двумя аллелями данного гена любой функции; один аллель может доминировать над вторым аллелем того же гена, рецессивным по отношению к третьему и совмещенным с четвертым. Кроме того, один аллель может быть доминирующим для одного признака, но не для других.
Доминирование - ключевое понятие в менделевской наследственности и классической генетике. Буквы и квадраты Пеннета используются для демонстрации принципов доминирования в обучении, а использование прописных букв для доминантных аллелей и строчных букв для рецессивных аллелей является широко применяемым соглашением. Классическим примером доминирования является наследование формы семян у гороха. Горох может быть круглым, ассоциированным с аллелем R, или морщинистым, ассоциированным с аллелем r. В этом случае возможны три комбинации аллелей (генотипов): RR, Rr и rr. RR (гомозиготные ) особи имеют круглый горошек, а rr (гомозиготные) особи - морщинистый горошек. У индивидов Rr (гетерозиготных ) аллель R маскирует присутствие аллеля r, поэтому у этих людей также есть круглый горошек. Таким образом, аллель R доминирует над аллелем r, а аллель r рецессивен по отношению к аллелю R.
Доминирование отличается от эпистаза, феномена аллеля одного гена, маскирующего влияние аллелей другой ген.
 Наследование карликовости кукурузы. Демонстрация роста растений двух родительских вариаций и их гетерозиготного гибрида F1 (в центре)
Наследование карликовости кукурузы. Демонстрация роста растений двух родительских вариаций и их гетерозиготного гибрида F1 (в центре) Концепция доминирования была введена Грегором Иоганном Менделем. Хотя Мендель, «отец генетики», впервые использовал этот термин в 1860-х годах, он не был широко известен до начала двадцатого века. Мендель заметил, что для множества признаков садового гороха, связанных с внешним видом семян, семенных коробочек и растений, существует два дискретных фенотипа, такие как круглые или морщинистые семена, желтые или зеленые семена, красные и белые цветы или высокие растения по сравнению с низкорослыми. При раздельном разведении растения всегда давали одни и те же фенотипы, поколение за поколением. Однако когда линии с разными фенотипами скрещивались (скрещивались), у потомства обнаруживался один и только один родительский фенотип (зеленый, круглый, красный или высокий). Однако, когда эти гибридные растения были скрещены, растения-потомки показали два исходных фенотипа в характерном соотношении 3: 1, причем наиболее распространенным фенотипом был фенотип родительских гибридных растений. Мендель рассуждал, что каждый родитель в первом скрещивании был гомозиготным по разным аллелям (один родительский AA и другой родительский aa), что каждый вносил один аллель в потомство, в результате чего все эти гибриды были гетерозиготами (Aa), и что один из двух аллелей в гибридном кроссе доминировал над экспрессией другого: A замаскированный a. Окончательное скрещивание двух гетерозигот (Aa X Aa) даст потомство AA, Aa и aa в соотношении генотипов 1: 2: 1, причем первые два класса демонстрируют фенотип (A), а последний - фенотип (a)., таким образом, обеспечивая соотношение фенотипов 3: 1.
Мендель не использовал термины ген, аллель, фенотип, генотип, гомозигота и гетерозигота, все из которых были введены позже. Он ввел обозначения прописных и строчных букв для доминантных и рецессивных аллелей, соответственно, которые используются до сих пор.
В 1928 году британский популяционный генетик Рональд Фишер предположил, что доминирование действует на основе естественного отбора за счет вклада генов-модификаторов. В 1929 году американский генетик Сьюэлл Райт ответил, заявив, что доминирование - это просто физиологическое следствие метаболических путей и относительной необходимости задействованного гена. Объяснение Райта стало установленным фактом в генетике, и дебаты в основном прекратились. Однако на некоторые черты могут влиять эволюционные механизмы.
Большинство животных и некоторые растения имеют парные хромосомы и описываются как диплоид. У них есть две версии каждой хромосомы, одна из которых внесена яйцеклеткой матери, а другая - сперматозоидом отца, известная как гаметы, описанная как гаплоидная, и создан посредством мейоза. Затем эти гаметы сливаются во время оплодотворения во время полового размножения в новую одноклеточную зиготу, которая многократно делится, в результате чего возникает новый организм с таким же количеством пары хромосом в каждой (негаметной) клетке в качестве ее родителей.
Каждая хромосома из совпадающей (гомологичной) пары структурно подобна другой и имеет очень похожую последовательность ДНК (локусов, сингулярный локус). ДНК в каждой хромосоме функционирует как серия дискретных генов, которые влияют на различные признаки. Таким образом, каждый ген также имеет соответствующий гомолог, который может существовать в разных версиях, называемых аллелями. Аллели одного и того же локуса на двух гомологичных хромосомах могут быть идентичными или разными.
группа крови человека определяется геном, который создает группу крови A, B, AB или O и расположен в длинном плече хромосомы. девять. Есть три разных аллеля, которые могут присутствовать в этом локусе, но только два могут присутствовать у любого человека: один унаследован от их матери, а другой - от отца.
Если два аллеля данного гена идентичны, организм называется гомозиготным и считается гомозиготным по этому гену; если вместо этого два аллеля различны, организм является гетерозиготным и гетерозиготным. Генетический состав организма, находящийся либо в одном локусе, либо по всем его генам вместе, называется его генотипом. Генотип организма прямо или косвенно влияет на его молекулярные, физические и другие характеристики, которые по отдельности или вместе называются его фенотипом. В гетерозиготных генных локусах эти два аллеля взаимодействуют, создавая фенотип.
При полном доминировании влияние одного аллеля в гетерозиготном генотипе полностью маскирует влияние другого. Аллель, который маскирует другой, считается доминантным для последнего, а маскируемый аллель считается рецессивным для первого. Следовательно, полное доминирование означает, что фенотип гетерозиготы неотличим от фенотипа доминирующей гомозиготы.
Классическим примером доминирования является наследование формы семян (формы гороха) у гороха. Горох может быть круглым (ассоциированный с аллелем R) или морщинистым (ассоциированным с аллелем r). В этом случае возможны три комбинации аллелей (генотипы ): RR и rr гомозиготны, а Rr гетерозиготны. У особей RR горошек круглый, у особей RR - морщинистый. У людей Rr аллель R маскирует присутствие аллеля r, поэтому у этих людей также есть круглый горошек. Таким образом, аллель R полностью доминантен по отношению к аллелю r, а аллель r рецессивен по отношению к аллелю R.
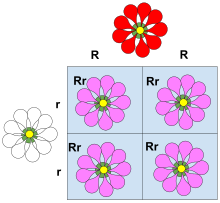 Этот квадрат Пеннета иллюстрирует неполное доминирование. В этом примере признак красного лепестка, связанный с аллелем R , рекомбинирует с признаком белого лепестка аллеля r. Растение не полностью выражает доминантный признак (R), вызывающий у растений с генотипом Rr экспрессию цветов с меньшим количеством красного пигмента, что приводит к появлению розовых цветов. Цвета не смешиваются друг с другом, доминантный признак просто выражен менее сильно.
Этот квадрат Пеннета иллюстрирует неполное доминирование. В этом примере признак красного лепестка, связанный с аллелем R , рекомбинирует с признаком белого лепестка аллеля r. Растение не полностью выражает доминантный признак (R), вызывающий у растений с генотипом Rr экспрессию цветов с меньшим количеством красного пигмента, что приводит к появлению розовых цветов. Цвета не смешиваются друг с другом, доминантный признак просто выражен менее сильно. Неполное доминирование (также называемое частичным доминированием, полудоминантным или промежуточным наследованием) возникает, когда фенотип гетерозиготного генотипа отличается от фенотипа гетерозиготного генотипа и часто является промежуточным с ним. фенотипы гомозиготных генотипов. Например, цвет цветка львиный зев гомозиготен по красному или белому цвету. Когда красный гомозиготный цветок сочетается с белым гомозиготным цветком, в результате получается розовый цветок львиный зев. Розовый львиный зев - результат неполного доминирования. Подобный тип неполного доминирования обнаруживается у четырехчасового растения, где розовый цвет получается при скрещивании чистокровных родителей белых и красных цветов. В количественной генетике, где фенотипы измеряются и обрабатываются численно, если фенотип гетерозиготы находится точно между (численно) фенотипом двух гомозигот, считается, что фенотип вообще не проявляет доминирования, т. Е. Доминирование существует только тогда, когда мера фенотипа гетерозиготы ближе к одной гомозиготе, чем к другой.
Когда растения поколения F 1 самоопыляются, фенотипическое и генотипическое соотношение поколения F 2 будет 1: 2: 1 (красный: Розовый: Белый).
См. гипотезу частичного доминирования.
 Содоминантность в Камелии сорта
Содоминантность в Камелии сорта  Группы крови A и B у людей демонстрируют совместное доминирование, но тип O рецессивен по отношению к A и B.
Группы крови A и B у людей демонстрируют совместное доминирование, но тип O рецессивен по отношению к A и B.  Этот квадрат Пеннета показывает совместное доминирование. В этом примере белый бык (WW) спаривается с красной коровой (RR), и их потомство демонстрирует совместное доминирование, выражающее как белые, так и рыжие волосы.
Этот квадрат Пеннета показывает совместное доминирование. В этом примере белый бык (WW) спаривается с красной коровой (RR), и их потомство демонстрирует совместное доминирование, выражающее как белые, так и рыжие волосы. Совместное доминирование возникает, когда видны вклады обоих аллелей. в фенотипе.
Например, в системе группы крови ABO химические модификации гликопротеина (антиген H) на поверхности клеток крови контролируются тремя аллелями, два из которых являются совмещенными друг с другом (I, I) и доминируют над рецессивным i в локусе ABO. Аллели I и I производят разные модификации. Фермент, кодируемый I, добавляет N-ацетилгалактозамин к мембраносвязанному H-антигену. Фермент I добавляет галактозу. Аллель i не вызывает модификации. Таким образом, каждый из аллелей I и I является доминантным для i (люди II и Ii оба имеют кровь типа A, а люди II и Ii оба имеют кровь типа B), но люди II имеют обе модификации в своих клетках крови и, таким образом, имеют кровь типа AB, поэтому аллели I и I считаются содоминантными.
Другой пример встречается в локусе для бета-глобинового компонента гемоглобина, где три молекулярных фенотипа: Hb / Hb, Hb / Hb и Hb / Все гемоглобины различимы с помощью электрофореза белков. (Медицинское состояние, вызванное гетерозиготным генотипом, называется серповидно-клеточным признаком и является более мягким состоянием, отличным от серповидно-клеточной анемии, таким образом, аллели демонстрируют неполное преобладание по отношению к анемии, см. выше). Для большинства локусов генов на молекулярном уровне оба аллеля экспрессируются совместно, потому что оба транскрибируются в РНК.
Ко-доминантность, при которой аллельные продукты сосуществуют в фенотипе, является отличается от неполного доминирования, когда количественное взаимодействие продуктов аллелей дает промежуточный фенотип. Например, при совместном доминировании красный гомозиготный цветок и белый гомозиготный цветок будут давать потомство с красными и белыми пятнами. Когда растения поколения F1 самоопыляются, фенотипическое и генотипическое соотношение поколения F2 будет 1: 2: 1 (красный: пятнистый: белый). Эти соотношения такие же, как и для неполного доминирования. Опять же, эта классическая терминология неуместна - на самом деле нельзя говорить о том, что такие случаи демонстрируют доминирование.
Хотя часто удобно говорить о рецессивном аллеле или доминантном признаке, доминирование не присуще ни аллелю, ни его фенотипу. Доминирование - это связь между двумя аллелями гена и их связанными фенотипами. «Доминантный» аллель является доминантным по отношению к конкретному аллелю того же гена, который может быть выведен из контекста, но он может быть рецессивным по отношению к третьему аллелю и кодоминантным по отношению к четвертому. Точно так же «рецессивный» признак - это признак, связанный с конкретным рецессивным аллелем, подразумеваемым контекстом, но тот же самый признак может встречаться в другом контексте, где он обусловлен каким-то другим геном и доминантным аллелем.
Доминирование не связано с природой самого фенотипа, то есть с тем, считается ли он «нормальным» или «ненормальным», «стандартным» или «нестандартным», «здоровым» или «больным» » сильнее »или« слабее »или более или менее экстремально. Доминантный или рецессивный аллель может объяснять любой из этих типов признаков.
Доминирование не определяет, является ли аллель вредоносным, нейтральным или полезным. Однако отбор должен опосредованно воздействовать на гены через фенотипы, а доминирование влияет на выявление аллелей в фенотипах и, следовательно, на скорость изменения частот аллелей при отборе. Вредные рецессивные аллели могут сохраняться в популяции с низкой частотой, при этом большинство копий переносятся в гетерозиготах бесплатно для этих людей. Эти редкие рецессивы являются основой многих наследственных генетических нарушений.
Доминирование также не связано с распределением аллелей в популяции. Как доминантные, так и рецессивные аллели могут быть чрезвычайно частыми или чрезвычайно редкими.
В генетике символы начинались как алгебраические заполнители. Когда один аллель является доминантным по отношению к другому, согласно старейшим правилам, доминантный аллель обозначается заглавной буквой. Рецессивному аллелю присваивается такая же буква в нижнем регистре. В примере с горохом, если известна взаимосвязь доминирования между двумя аллелями, можно обозначить доминантный аллель, который дает округлую форму, заглавным символом R, а рецессивный аллель, который дает морщинистая форма строчными буквами r . Гомозиготный доминантный, гетерозиготный и гомозиготный рецессивный генотипы обозначаются как RR, Rrи rr соответственно. Также можно было бы обозначить два аллеля как W и w, а также три генотипа WW, Wwи ww, первые два из которых дали горошек круглый и третий горошек морщинистый. Выбор «R » или «W » в качестве символа доминантного аллеля не предопределяет, является ли аллель, вызывающий «круглый» или «морщинистый» фенотип, когда гомозиготен. доминирующий.
У гена может быть несколько аллелей. Каждый аллель обозначен символом локуса, за которым следует уникальный верхний индекс. У многих видов самый распространенный аллель в дикой популяции обозначается как аллель дикого типа. Он обозначается знаком + в качестве надстрочного индекса. Другие аллели являются доминантными или рецессивными по отношению к аллелю дикого типа. Для рецессивных аллелей символ локуса пишется строчными буквами. Для аллелей с любой степенью доминирования по отношению к аллелю дикого типа первая буква символа локуса находится в верхнем регистре. Например, вот некоторые из аллелей в локусе a лабораторной мыши Mus musculus: A, доминантный желтый; а - дикий тип; и а, черно-подпалый. Аллель a является рецессивным по отношению к аллелю дикого типа, а аллель A кодоминантен аллелю дикого типа. Аллель A также кодоминантен аллелю a, но показывает, что это отношение выходит за рамки правил генетической номенклатуры мышей.
Правила генетической номенклатуры эволюционировали, поскольку генетика стала более сложной. Комитеты стандартизировали правила для некоторых видов, но не для всех. Правила для одного вида могут несколько отличаться от правил для другого вида.
Хотя любой особь диплоидного организма имеет не более два разных аллеля в любом локусе (за исключением анеуплоидий ), большинство генов существует в большом количестве аллельных версий в популяции в целом. Если аллели по-разному влияют на фенотип, иногда их отношения доминирования можно описать как серию.
Например, на цвет шерсти домашних кошек влияет серия аллелей гена TYR (который кодирует фермент тирозиназу ). Аллели C, c, c и c (полноцветный, бирманский, сиамский и альбинос, соответственно) продуцируют разные уровни пигмента и, следовательно, разные уровни разбавления цвета. Аллель C (полный цвет) полностью доминирует над последними тремя, а аллель c (альбинос) полностью рецессивен по отношению к первым трем.
У людей и у других млекопитающих видов, пол определяется двумя половыми хромосомами, называемыми Х-хромосомой и Y-хромосомой. Человеческие женщины обычно XX ; мужчины обычно XY . Остальные пары хромосом встречаются у обоих полов и называются аутосомами ; генетические признаки, обусловленные локусами на этих хромосомах, описываются как аутосомные и могут быть доминантными или рецессивными. Генетические признаки на хромосомах X и Y называются сцепленными с полом, потому что они связаны с половыми хромосомами, а не потому, что они характерны для одного пола или другого. На практике этот термин почти всегда относится к чертам, связанным с X, и на очень многие такие черты (например, недостаточность цветового зрения в красно-зеленых тонах) пол не влияет. У женщин есть две копии каждого локуса гена, обнаруженного на Х-хромосоме, так же, как и для аутосом, и применяются те же отношения доминирования. Однако самцы имеют только одну копию каждого локуса гена Х-хромосомы и описываются как гемизиготные для этих генов. Y-хромосома намного меньше, чем X, и содержит гораздо меньший набор генов, включая, помимо прочего, те, которые влияют на «мужественность», такие как SRY ген фактора, определяющего семенник. Правила доминирования локусов сцепленных с полом генов определяются их поведением у самки: поскольку у самца есть только один аллель (за исключением некоторых типов Y-хромосомы анеуплоидии ), этот аллель всегда выражается независимо от от того, является ли он доминантным или рецессивным. Птицы имеют противоположные половые хромосомы: у самцов - ZZ, у самок - ZW. Однако в остальном наследование признаков напоминает XY-систему; самцы зебровых зябликов могут нести ген белой окраски в одной из двух Z-хромосом, но у самок всегда белая окраска. У кузнечиков есть XO-система. У женщин XX, а у мужчин только X. Y-хромосомы вообще нет.
Эпистаз ["epi + stasis = сидеть на вершине"] - это взаимодействие между аллелями в двух разных локусах генов, которые влияют на один признак, что иногда может напоминать доминирующее взаимодействие между два разных аллеля в одном локусе. Эпистаз изменяет характеристическое соотношение 9: 3: 3: 1, ожидаемое для двух неэпистатических генов. Для двух локусов распознается 14 классов эпистатических взаимодействий. В качестве примера рецессивного эпистаза один локус гена может определять, является ли пигмент цветка желтым (AA или Aa ) или зеленым (aa ), в то время как другой локус определяет, продуцируется ли пигмент (BB или Bb ) или нет (bb ). У растения bb цветы будут белыми, независимо от генотипа другого локуса, например, AA, Aaили aa . Комбинация bb не является доминантной по отношению к аллелю A : скорее, ген B демонстрирует рецессивный эпистаз к A, поскольку локус B, когда он гомозиготен по рецессивному аллелю (bb ), подавляет фенотипическую экспрессию локуса A . При скрещивании двух растений AaBb это дает характерное соотношение 9: 3: 4, в данном случае желтые: зеленые: белые цветы.
В доминантном эпистазе один локус гена может определять желтый или зеленый пигмент, как в предыдущем примере: AA и Aa желтые, и aa зеленые. Второй локус определяет, продуцируется ли предшественник пигмента (dd ) или нет (DD или Dd ). Здесь, у растения DD или Dd, цветы будут бесцветными независимо от генотипа в локусе A из-за эпистатического эффекта доминантного D аллель. Таким образом, при скрещивании двух растений AaDd 3/4 растений будут бесцветными, и фенотипы желтого и зеленого цвета экспрессируются только у растений dd . Это дает характерное соотношение 12: 3: 1 белых: желтых: зеленых растений.
Дополнительный эпистаз возникает, когда два локуса влияют на один и тот же фенотип. Например, если цвет пигмента создается с помощью CC или Cc, но не cc, и с помощью DD или Dd, но не dd, то пигмент не продуцируется ни в одной генотипической комбинации с cc или dd . То есть оба локуса должны иметь по крайней мере один доминантный аллель для создания фенотипа. Это дает характерное соотношение 9: 7 пигментированных и непигментированных растений. Комплементарный эпистаз, напротив, дает непигментированное растение тогда и только тогда, когда генотип cc и dd, а характеристическое соотношение составляет 15: 1 между пигментированными и непигментированными растениями.
Классическая генетика рассматривала эпистатические взаимодействия между двумя генами одновременно. В настоящее время из молекулярной генетики очевидно, что все локусы генов вовлечены в сложные взаимодействия со многими другими генами (например, метаболические пути могут включать множество генов), и что это создает эпистатические взаимодействия, которые намного сложнее, чем в классических двухлокусных моделях..
Частоту гетерозиготного состояния (которое является состоянием носителя рецессивного признака) можно оценить с помощью Харди – Вайнберга формула :
Эта формула применяется к гену с ровно двумя аллелями и связывает частоты этих аллелей в большой популяции с частотами их трех генотипов в этой популяции.
Например, если p - частота аллеля A, а q - частота аллеля a, то термины p, 2pq и q - частоты генотипов AA, Aaи aa соответственно. Поскольку ген имеет только два аллеля, все аллели должны быть либо A, либо a и p + q = 1. Теперь, если A полностью доминантен для a, то частоту генотипа носителя Aa нельзя непосредственно наблюдать (поскольку он имеет те же признаки, что и гомозиготный генотип AA ), однако ее можно оценить из частота рецессивного признака в популяции, поскольку она такая же, как и у гомозиготного генотипа aa . т.е. частоту отдельных аллелей можно оценить: q = √ f(aa), p = 1 - q, и из них можно определить частоту генотипа носителя: f( Аа) = 2пк.
Эта формула основывается на ряде допущений и точной оценке частоты рецессивного признака. В общем, любая реальная ситуация будет в некоторой степени отклоняться от этих предположений, внося соответствующие неточности в оценку. Если рецессивный признак встречается редко, то будет трудно точно оценить его частоту, так как потребуется очень большой размер выборки.
Свойство «доминантный» иногда путают с понятием «выгодный», а свойство «рецессивный» иногда путают с понятием «вредный», но явления различны. Доминирование описывает фенотип гетерозигот в отношении фенотипов гомозигот и независимо от того, в какой степени различные фенотипы могут быть полезными или вредными. Поскольку многие аллели генетических заболеваний рецессивны и слово «доминирование» имеет положительный оттенок, часто делается предположение, что доминирующий фенотип превосходит приспособленность. Однако это не гарантируется; как обсуждается ниже, хотя большинство аллелей генетических заболеваний являются вредными и рецессивными, не все генетические заболевания являются рецессивными.
Тем не менее, эта путаница была повсеместной на протяжении всей истории генетики и сохраняется по сей день. Устранение этой путаницы было одним из главных мотивов для публикации принципа Харди – Вайнберга.
Мендель не знал молекулярных основ доминирования. Теперь понятно, что локус гена включает длинный ряд (от сотен до тысяч) из оснований или нуклеотидов дезоксирибонуклеиновой кислоты (ДНК) в определенной точке на хромосома. центральная догма молекулярной биологии гласит, что «ДНК создает РНК создает белок », то есть ДНК транскрибируется для создания копии РНК, а РНК транслируется для создания белка. В этом процессе разные аллели в локусе могут транскрибироваться, а могут и не транскрибироваться, и в случае транскрибирования могут транслироваться в несколько разные версии одного и того же белка (так называемые изоформы ). Белки часто действуют как ферменты, которые катализируют химические реакции в клетке, прямо или косвенно производящие фенотипы. В любом диплоидном организме последовательности ДНК двух аллелей, присутствующих в любом локусе гена, могут быть идентичными (гомозиготными) или разными (гетерозиготными). Даже если локус гена гетерозиготен на уровне последовательности ДНК, белки, производимые каждым аллелем, могут быть идентичными. При отсутствии каких-либо различий между белковыми продуктами ни один из аллелей не может считаться доминантным (см. Совместное доминирование выше). Даже если два белковых продукта немного отличаются (аллозимы ), вполне вероятно, что они продуцируют один и тот же фенотип в отношении действия фермента, и снова ни один аллель не может считаться доминантным.
Доминирование обычно происходит, когда один из двух аллелей нефункционален на молекулярном уровне, то есть не транскрибируется или не производит функциональный белок товар. Это может быть результатом мутации , которая изменяет последовательность ДНК аллеля. Организм, гомозиготный по нефункциональному аллелю, обычно будет демонстрировать отличительный фенотип из-за отсутствия белкового продукта. Например, у людей и других организмов непигментированная кожа фенотипа альбинос возникает, когда человек гомозиготен по аллелю, который кодирует нефункциональную версию фермента, необходимого для производства пигмента кожи меланин. Важно понимать, что не отсутствие функции позволяет описывать аллель как рецессивный: это взаимодействие с альтернативным аллелем в гетерозиготе. Возможны три основных типа взаимодействия:
Многие белки обычно активны в форме мультимера, совокупность нескольких копий одного и того же белка, иначе известная как гомомультимерный белок или гомоолигомерный белок. Фактически, большинство из 83 000 различных ферментов из 9800 различных организмов в базе данных ферментов BRENDA представляют собой гомоолигомеры. Когда версия белка дикого типа присутствует вместе с мутантной версией, может быть образован смешанный мультимер. Мутация, которая приводит к появлению мутантного белка, который нарушает активность белка дикого типа в мультимере, является доминантно-отрицательной мутацией.
Доминантно-отрицательная мутация может возникнуть в соматической клетке человека и обеспечить пролиферативное преимущество мутантной клетки, что приведет к ее клональной экспансии. Например, доминантно-отрицательная мутация в гене, необходимая для нормального процесса запрограммированной гибели клеток (апоптоз ) в ответ на повреждение ДНК, может сделать клетку устойчивой к апоптозу. Это позволит клону размножаться даже при чрезмерном повреждении ДНК. Такие доминантно-отрицательные мутации возникают в гене-супрессоре опухоли p53. Белок P53 дикого типа обычно присутствует в виде четырехбелкового мультимера (олиготетрамера). Доминантно-отрицательные мутации р53 встречаются при различных типах рака и предраковых поражениях (например, опухолях мозга, раке груди, предраковых поражениях полости рта и раке полости рта).
Доминантно-отрицательные мутации также встречаются в другие гены-супрессоры опухолей. Например, две доминантно-отрицательные мутации зародышевой линии были идентифицированы в гене , мутировавшем телеангиэктазию Ataxia (ATM), который увеличивает восприимчивость к раку груди. Доминирующие негативные мутации фактора транскрипции C / EBPα могут вызывать острый миелоидный лейкоз. Унаследованные доминантно-отрицательные мутации также могут повышать риск других заболеваний, помимо рака. Доминантно-отрицательные мутации в рецепторе, активируемом пролифератором пероксисом (PPARγ), связаны с тяжелой инсулинорезистентностью, сахарным диабетом и гипертонией.
люди. Фактически, первое исследование, сообщающее о мутантном белке, ингибирующем нормальную функцию белка дикого типа в смешанном мультимере, было с белком хвостового волокна бактериофага Т4 GP37. Мутации, которые производят усеченный белок, а не полноразмерный мутантный белок, по-видимому, имеют самый сильный доминантно-негативный эффект в исследованиях P53, ATM, C / EBPα и бактериофага T4 GP37.
Многие генетические признаки или заболевания человека классифицируются просто как «доминантные» или «рецессивные». Особенно с так называемыми рецессивными заболеваниями, которые действительно являются фактором рецессивных генов, но могут чрезмерно упрощать лежащие в основе m на молекулярной основе и приводят к непониманию природы доминирования. Например, рецессивное генетическое заболевание фенилкетонурия (PKU) является результатом любого из большого числа (>60) аллелей в локусе гена для фермента фенилаланингидроксилазы (PAH ). Многие из этих аллелей производят мало или не производят PAH, в результате чего субстрат фенилаланин (Phe) и его побочные продукты метаболизма накапливаются в центральной нервной системе и может вызвать серьезную умственную отсталость при отсутствии лечения.
Чтобы проиллюстрировать эти нюансы, генотипы и фенотипические последствия взаимодействий между тремя гипотетическими аллелями PAH показаны в следующей таблице:
| Генотип | PAH активность | [Phe ] конц | ФКУ? |
|---|---|---|---|
| AA | 100% | 60 мкМ | Нет |
| AB | 30% | 120 мкМ | Нет |
| CC | 5% | 200 ~ 300 мкМ | Гиперфенилаланинемия |
| BB | 0,3% | 600 ~ 2400 мкМ | Да |
У здоровых людей, гомозиготных по стандартному функциональному аллелю (AA ), активность PAH является стандартной (100%), а концентрация фенилаланина в крови [Phe ] составляет около 60 мкМ (= мкмоль / л ). У нелеченных лиц, гомозиготных по одному из аллелей PKU (BB ), активность PAH близка к нулю, [Phe] в десять-сорок раз превышает стандартную, и индивидуум проявляет PKU.
В гетерозиготе AB активность PAH составляет только 30% (а не 50%) от стандарта, в крови [Phe ] повышено на два -кратно, и у человека не проявляется ФКУ. Таким образом, аллель A является доминантным по отношению к аллелю B в отношении PKU, но аллель B не полностью доминирует по отношению к аллелю A аллель в отношении его молекулярного эффекта, определение уровня активности PAH (0,3% < 30% << 100%). Finally, the A аллель является неполным доминантным для B по отношению к [Phe], поскольку 60 мкМ < 120 μM << 600 μM. Note once more that it is irrelevant to the question of dominance that the recessive allele produces a more extreme [Phe] phenotype.
Для третьего аллеля C гомозигота CC продуцирует очень небольшое количество фермента PAH, что приводит к несколько повышенному уровню [ Phe ] в крови, состояние, называемое гиперфенилаланинемия, которое не приводит к умственной отсталости.
То есть отношения доминирования любых двух аллелей могут варьироваться в зависимости от какой аспект фенотипа рассматривается. Обычно более полезно говорить о фенотипических последствиях аллельных взаимодействий, вовлеченных в любой генотип, чем пытаться разделить их на доминантные и рецессивные категории.
