Войти
| Просмотр / редактирование человека | Просмотр / редактирование мыши |
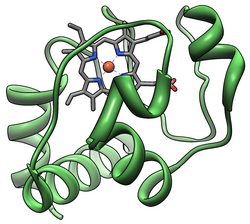
 Простетическая группа гема цитохрома c, состоящая из жесткого порфиринового кольца, координированного с атомом железа.
Простетическая группа гема цитохрома c, состоящая из жесткого порфиринового кольца, координированного с атомом железа. цитохромный комплекс, или cyt c, представляет собой небольшой гемепротеин, который обнаруживается, свободно ассоциированный с внутренней мембраной митохондрии. Он принадлежит к белкам семейства цитохрома c и играет важную роль в апоптозе клеток. Цитохром с хорошо растворим в воде, в отличие от других цитохромов, и является важным компонентом цепи переноса электронов, где он несет один электрон. Он способен подвергаться окислению и восстановлению, поскольку его атом железа превращается между формами двухвалентного железа и трехвалентного железа, но не связывает кислород. Он переносит электроны между комплексами III (кофермент Q - Cyt C редуктаза) и IV (Cyt C оксидаза). У человека цитохром c кодируется геном CYCS .
Цитохром c - это высококонсервативный белок для всего спектра видов, обнаруженный у растений, животных и многих одноклеточных организмов. Это, наряду с его небольшим размером (молекулярная масса около 12000 дальтон ), делает его полезным в исследованиях кладистики. Молекула цитохрома с изучалась с точки зрения эволюционной биологии.
Цитохром c имеет первичную структуру, состоящую из цепи примерно из 100 аминокислот. Многие организмы высшего порядка обладают цепочкой из 104 аминокислот. Последовательности цитохрома c у людей идентичны последовательностям шимпанзе (наши ближайшие родственники), но отличаются от последовательностей лошадей.
Цитохром c имеет аминокислотную последовательность, которая является высококонсервативной у эукариот и отличается только одним немного остатков. У более чем тридцати видов, протестированных в одном исследовании, 34 из 104 аминокислот были консервативными; идентичны в их характерном положении. Например, человеческая цитохромоксидаза реагирует с цитохромом с пшеницы in vitro; что справедливо для всех пар исследованных видов. Кроме того, окислительно-восстановительный потенциал +0,25 В одинаков для всех исследованных молекул цитохрома c.
 Кристаллы цитохрома c Тунафиш (длиной ~ 5 мм), выращенные путем диффузии жидкость-жидкость в условиях микрогравитации в
Кристаллы цитохрома c Тунафиш (длиной ~ 5 мм), выращенные путем диффузии жидкость-жидкость в условиях микрогравитации в Цитохром c принадлежит к классу I семейства цитохромов c-типа и содержит характерный аминокислотный мотив CXXCH (цистеин-любой-любой-цистеин-гистидин), который связывает гем.. Это мотив расположен по направлению к N-концу цепи пептида и содержит гистидин в качестве пятого лиганда гемового железа. Шестой лиганд представлен остатком метионина, обнаруженным у С-конца. Основа белка свернута в пять α-спиралей, пронумерованных α1-α5 от N-конца до C-конца. Спирали α3, α4 и α5 называются спиралями 50, 60 и 70 соответственно при обращении к митохондриальному цитохрому c.
 Структура гема c
Структура гема c В то время как большинство гемовых белков прикреплены к простетическая группа посредством лигирования ионов железа и третичных взаимодействий, гемовая группа цитохрома с образует тиоэфирные связи с двумя боковыми цепями цистеина белка. Одним из основных свойств гема с, который позволяет цитохрому с выполнять множество функций, является его способность иметь в природе различные восстановительные потенциалы. Это свойство определяет кинетику и термодинамику реакции переноса электрона.
Дипольный момент играет важную роль в ориентации белков в правильном направлении и повышении их способности связываться с другими молекулами.. Дипольный момент цитохрома с является результатом кластера отрицательно заряженных боковых цепей аминокислот в «задней части» фермента. Несмотря на вариации в количестве связанных гемовых групп и вариации в последовательности, дипольный момент цитохромов c позвоночных заметно сохраняется. Например, все цитохромы c позвоночных имеют дипольный момент приблизительно 320 дебай, в то время как цитохромы c растений и насекомых имеют дипольный момент приблизительно 340 дебай.
Цитохром c является компонентом цепи переноса электронов в митохондриях. Группа гема цитохрома c принимает электроны от bc1комплекса и переносит электроны в комплекс IV. Цитохром c также участвует в инициации апоптоза. При высвобождении цитохрома c в цитоплазму белок связывает фактор-1, активирующий апоптотическую протеазу (Apaf-1).
Цитохром c может также катализировать несколько окислительно-восстановительных реакций, таких как гидроксилирование и ароматическое окисление и проявляет пероксидазную активность за счет окисления различных доноров электронов, таких как 2,2-азино-бис (3-этилбензтиазолин-6-сульфоновая кислота) (ABTS ), 2-кето-4-тиометилмасляная кислота и 4-аминоантипирин.
Бактериальный цитохром с функционирует как нитритредуктаза.
Цитохром с также был обнаружен в 1996 году доктором. Сяодун Ван играет промежуточную роль в апоптозе, контролируемой форме гибели клеток, используемой для уничтожения клеток в процессе развития или в ответ на инфекцию или повреждение ДНК.
Цитохром. c связывается с кардиолипином во внутренней митохондриальной мембране, тем самым закрепляя его присутствие и не позволяя ему высвобождаться из митохондрий и инициировать апоптоз. В то время как первоначальное притяжение между кардиолипином и цитохромом c является электростатическим из-за чрезвычайно положительного заряда цитохрома c, конечное взаимодействие является гидрофобным, когда гидрофобный хвост кардиолипина вставляется в гидрофобную часть цитохрома c.
На ранней стадии апоптоза продукция митохондриальных АФК стимулируется, а кардиолипин окисляется пероксидазной функцией комплекса кардиолипин-цитохром с. Затем гемопротеин отделяется от внутренней митохондриальной мембраны и может быть экструдирован в растворимую цитоплазму через поры на внешней мембране.
Устойчивое повышение уровня кальция предшествует высвобождению цит с из митохондрий.. Высвобождение небольших количеств cyt c приводит к взаимодействию с рецептором IP3 (IP3R) на эндоплазматическом ретикулуме (ER), вызывая высвобождение кальция ER. Общее увеличение кальция запускает массивное высвобождение cyt c, который затем действует в петле положительной обратной связи, поддерживая высвобождение кальция ER через IP3R. Это объясняет, как высвобождение кальция из ER может достигать цитотоксических уровней. Это высвобождение цитохрома с, в свою очередь, активирует каспазу 9, цистеиновую протеазу. Затем каспаза 9 может активировать каспазу 3 и каспазу 7, которые отвечают за разрушение клетки изнутри.
Одним из способов активации апоптоза клеток является высвобождение цитохрома с из митохондрий в цитозоль. Исследование показало, что клетки способны защищать себя от апоптоза, блокируя высвобождение цитохрома с с помощью Bcl-x L. Другой способ, которым клетки могут контролировать апоптоз, - это фосфорилирование Tyr48, которое превратит цитохром c в антиапоптотический переключатель.
 Удаление O и H 2O2цитохромом c
Удаление O и H 2O2цитохромом c Известно, что цитохром c играет роль в цепи переноса электронов и апоптозе клетки. Однако недавнее исследование показало, что он также может действовать как антиоксидантный фермент в митохондриях; и это достигается путем удаления супероксида (O2) и перекиси водорода (H2O2) из митохондрий. Следовательно, цитохром c необходим не только в митохондриях для клеточного дыхания, но он также необходим в митохондриях для ограничения продукции O 2 и H 2O2.
Цитохром Считается, что c локализуется исключительно в митохондриальном межмембранном пространстве при нормальных физиологических условиях. Высвобождение цитохрома-с из митохондрий в цитозоль, где он активирует семейство каспаз из протеаз, считается основным триггером, приводящим к началу апоптоза. Измерение количества цитохрома с, просачивающегося из митохондрий в цитозоль и из клетки в культуральную среду, является чувствительным методом контроля степени апоптоза. Однако подробные иммуноэлектронные микроскопические исследования срезов тканей крысы с использованием специфичных к цитохрому с антител предоставляют убедительные доказательства того, что цитохром-с в нормальных клеточных условиях также присутствует во внемитохондриальных участках. В ацинарных клетках поджелудочной железы и передней доли гипофиза сильное и специфическое присутствие цитохрома-c было обнаружено в гранулах зимогена и в гранулах гормона роста соответственно. В поджелудочной железе цитохром-с также был обнаружен в конденсирующихся вакуолях и в ацинарном просвете. Было показано, что экстрамитохондриальная локализация цитохрома c является специфической, поскольку она полностью исчезает при адсорбции первичного антитела очищенным цитохромом c. Присутствие цитохрома-с вне митохондрий в определенном месте при нормальных физиологических условиях поднимает важные вопросы, касающиеся его клеточной функции и механизма транслокации. Помимо цитохрома с, внемитохондриальная локализация также наблюдалась для большого количества других белков, включая те, которые кодируются митохондриальной ДНК. Это повышает вероятность существования пока неустановленных специфических механизмов транслокации белков из митохондрий в другие клеточные места назначения.
 Пероксинизотистая кислота
Пероксинизотистая кислота Был использован цитохром c для обнаружения производства перекиси в биологических системах. По мере образования супероксида количество окисленного цитохрома с увеличивается, а восстановленного цитохрома с уменьшается. Однако супероксид часто получают из оксида азота. В присутствии оксида азота восстановление цитохрома с подавляется. Это приводит к окислению цитохрома с до цитохрома с пероксизотистой кислотой, промежуточным продуктом, образующимся в результате реакции оксида азота и супероксида. Присутствие в митохондриях пероксинитрита или H 2O2и диоксида азота NO2может быть летальным, поскольку они нитрируют тирозин остатки цитохрома с, что приводит к нарушению цитохрома с. функционируют как переносчик электронов в цепи переноса электронов.
| Wikimedia Commons имеет среду, относящуюся к цитохрому c. |