
Войти
| Спайк гликопротеина S1 коронавируса | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Модель внешнего строения вириона SARS-CoV-2. ● Синий: конверт ● Бирюзовый: шип гликопротеина (S) ● Красный: белки оболочки (E) ● Зеленый: мембранные белки (M) ● Оранжевый: гликан Модель внешнего строения вириона SARS-CoV-2. ● Синий: конверт ● Бирюзовый: шип гликопротеина (S) ● Красный: белки оболочки (E) ● Зеленый: мембранные белки (M) ● Оранжевый: гликан | |||||||||
| Идентификаторы | |||||||||
| Условное обозначение | CoV_S1 | ||||||||
| Pfam | PF01600 | ||||||||
| ИнтерПро | IPR002551 | ||||||||
| |||||||||
| Спайк гликопротеина S2 коронавируса | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||
| Условное обозначение | CoV_S2 | ||||||||
| Pfam | PF01601 | ||||||||
| ИнтерПро | IPR002552 | ||||||||
| |||||||||
| Бетакоронавирусный гликопротеин S1, связывание с рецептором | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||
| Условное обозначение | bCoV_S1_RBD | ||||||||
| Pfam | PF09408 | ||||||||
| ИнтерПро | IPR018548 | ||||||||
| |||||||||
| Бетакоронавирусоподобный спайковый гликопротеин S1, N-концевой | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||
| Условное обозначение | bCoV_S1_N | ||||||||
| Pfam | PF16451 | ||||||||
| ИнтерПро | IPR032500 | ||||||||
| |||||||||
Белок Spike (S) (иногда также гликопротеин Spike , ранее известный как E2) является крупнейшим из четырех основных структурных белков, обнаруженных в коронавирусах. Белок-шип собирается в тримеры, которые образуют большие структуры, называемые шипами или пепломерами, которые выступают с поверхности вириона. Характерный внешний вид этих шипов при визуализации с помощью просвечивающей электронной микроскопии с негативным окрашиванием, «напоминающий солнечную корону », дал название семейству вирусов.
Функция белка-шипа состоит в том, чтобы опосредовать проникновение вируса в клетку-хозяин, сначала взаимодействуя с молекулами на внешней поверхности клетки, а затем сливая вирусную и клеточную мембраны. Спайковый белок - это гибридный белок класса I, который содержит две области, известные как S1 и S2, отвечающие за эти две функции. Область S1 содержит рецептор-связывающий домен, который связывается с рецепторами на поверхности клетки. Коронавирусы используют самые разные рецепторы; SARS-CoV (вызывающий SARS ) и SARS-CoV-2 (вызывающий COVID-19 ) взаимодействуют с ангиотензин-превращающим ферментом 2 (ACE2). Область S2 содержит слитый пептид и другую инфраструктуру слияния, необходимую для слияния мембраны с клеткой-хозяином, что является необходимым этапом для инфицирования и репликации вируса. Спайковый белок определяет круг хозяев вируса (какие организмы он может заразить) и клеточный тропизм (какие клетки или ткани он может инфицировать в организме).
Спайковый белок обладает высокой иммуногенностью. Антитела против белка спайков обнаружены у пациентов, переболевших SARS и COVID-19. Нейтрализующие антитела нацелены на эпитопы рецептор-связывающего домена. Большинство усилий по разработке вакцины против COVID-19 в ответ на пандемию COVID-19 нацелены на активацию иммунной системы против шипового белка.
 Структура тримерного белка шипа SARS-CoV-2 в конформации до слияния с криоэлектронной микроскопией с выделенным единственным мономером. S1 NTD показан синим, а S1 CTD (который служит рецептор-связывающим доменом) показан розовым. Спирали показывают оранжевым и голубым цветом части S2, которые претерпевают конформационные изменения во время слияния. Черная полоса внизу указывает положение вирусной мембраны. Из PDB : 6VSB.
Структура тримерного белка шипа SARS-CoV-2 в конформации до слияния с криоэлектронной микроскопией с выделенным единственным мономером. S1 NTD показан синим, а S1 CTD (который служит рецептор-связывающим доменом) показан розовым. Спирали показывают оранжевым и голубым цветом части S2, которые претерпевают конформационные изменения во время слияния. Черная полоса внизу указывает положение вирусной мембраны. Из PDB : 6VSB. Белок-спайк очень большой, часто длиной 1200-1400 аминокислотных остатков; в SARS-CoV-2 это 1273 остатка. Это однопроходный трансмембранный белок с коротким C-концевым хвостом внутри вируса, трансмембранной спиралью и большим N-концевым эктодоменом, выставленным снаружи вируса.
Белок-спайк образует гомотримеры, в которых три копии белка взаимодействуют через свои эктодомены. Структуры тримера были описаны как имеющие форму булавы или лепестка. Каждый спайковый белок содержит две области, известные как S1 и S2, и в собранном тримере области S1 на N-конце образуют часть белка, наиболее удаленную от вирусной поверхности, в то время как области S2 образуют гибкий «стержень», содержащий большую часть что белок-белковые взаимодействия, которые удерживают тример на месте.
Область S1 белка-шипа отвечает за взаимодействие с рецепторными молекулами на поверхности клетки-хозяина на первом этапе проникновения вируса. S1 содержит два домена, называемых N-концевым доменом (NTD) и C-концевым доменом (CTD), иногда также называемыми доменами A и B. В зависимости от коронавируса один или оба домена могут использоваться в качестве рецептор-связывающих доменов (RBD). Рецепторы-мишени могут быть самыми разнообразными, включая белки рецепторов клеточной поверхности и сахара, такие как сиаловые кислоты в качестве рецепторов или корецепторов. В общем, NTD связывает молекулы сахара, в то время как CTD связывает белки, за исключением вируса гепатита мышей, который использует свои NTD для взаимодействия с рецептором белка CEACAM1. НТД имеет Galectin -подобных белков раза, но молекулы сахара связывается несколько иначе, чем galectins.
CTD отвечает за взаимодействия MERS-CoV с его рецепторной дипептидилпептидазой-4, а также SARS-CoV и SARS-CoV-2 с их рецепторным ангиотензинпревращающим ферментом 2 (ACE2). CTD этих вирусов можно далее разделить на два субдомена, известных как ядро и расширенная петля или рецептор-связывающий мотив (RBM), где находится большинство остатков, которые непосредственно контактируют с рецептором-мишенью. Существуют тонкие различия, в основном в RBM, между взаимодействиями белков-шипов SARS-CoV и SARS-CoV-2 с ACE2. Сравнение белков-шипов из нескольких коронавирусов позволяет предположить, что дивергенция в области RBM может объяснять различия в рецепторах-мишенях, даже если ядро S1 CTD структурно очень похоже.
В клонах коронавирусов, а также в четырех основных подгруппах коронавирусов область S1 менее консервативна, чем S2, что соответствует ее роли во взаимодействии с вирус-специфическими рецепторами клетки-хозяина. В области S1 NTD более консервативна, чем CTD.
Область S2 белка-шипа отвечает за слияние мембран между вирусной оболочкой и клеткой-хозяином, обеспечивая проникновение генома вируса в клетку. Область S2 содержит гибридный пептид, участок, состоящий в основном из гидрофобных аминокислот, функция которого заключается в проникновении и дестабилизации мембраны клетки-хозяина. S2 также содержит два субдомена гептадного повтора, известные как HR1 и HR2, иногда называемые областью «ядра слияния». Эти субдомены претерпевают существенные конформационные изменения в процессе сварки с образованием шести спирали пучок, характерную особенность из слитых белков класса I. Также считается, что область S2 включает трансмембранную спираль и С-концевой хвост, расположенный внутри вириона.
По сравнению с S1, область S2 очень хорошо консервативна среди коронавирусов.
 Спайковый белок с гликозилированием и без него.
Спайковый белок с гликозилированием и без него. Спайковый белок представляет собой гликопротеин и сильно гликозилирован посредством N-связанного гликозилирования. Исследования шипового белка SARS-CoV-2 также сообщили о O-связанном гликозилировании в области S1. С-концевой хвост, расположенный внутри вириона, обогащен остатками цистеина и пальмитоилирован.
Спайковые белки активируются посредством протеолитического расщепления. Они расщепляются протеазами клетки-хозяина на границе S1-S2, а затем в так называемом сайте S2 'на N-конце гибридного пептида.
Как и другие слитые белки класса I, спайковый белок претерпевает очень большие конформационные изменения в процессе слияния. Состояния до и после слияния некоторых коронавирусов, особенно SARS-CoV-2, были изучены с помощью криоэлектронной микроскопии. Функционально важная динамика белка также наблюдалась в состоянии до слияния, в котором относительная ориентация некоторых из областей S1 относительно S2 в тримере может изменяться. В закрытом состоянии все три области S1 плотно упакованы, и область, которая контактирует с рецепторами клетки-хозяина, стерически недоступна, в то время как в открытых состояниях один или два RBD S1 более доступны для связывания рецептора в открытой или «верхней» конформации..
 Трансмиссионный электронный микроскоп из ТОРСА-COV-2 вириона, показывающий характеристический «Корона» внешний вид с острием белками (зелеными), образующий известные проекции с поверхности вириона (желтым).
Трансмиссионный электронный микроскоп из ТОРСА-COV-2 вириона, показывающий характеристический «Корона» внешний вид с острием белками (зелеными), образующий известные проекции с поверхности вириона (желтым). 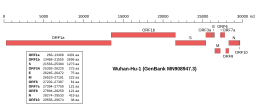 Геномная организация изолята Wuhan-Hu-1, самого раннего секвенированного образца SARS-CoV-2, указывающая на расположение гена S. Геномная организация изолята Wuhan-Hu-1, самого раннего секвенированного образца SARS-CoV-2, указывающая на расположение гена S. | |
| Идентификатор генома NCBI | 86693 |
|---|---|
| Размер генома | 29,903 базиса |
| Год окончания | 2020 г. |
| Браузер генома ( UCSC ) | |
Ген, кодирующий белок шип расположен по направлению к 3' - конце вируса в положительной смысловой РНК генома, наряду с генами трех других структурных белков и различных вирус-специфических вспомогательных белков. Транспортировка белков спайк-белков, по-видимому, зависит от подгруппы коронавирусов: при изолированной экспрессии без других вирусных белков, спайковые белки из бета-коронавирусов способны достигать клеточной поверхности, в то время как белки из альфа- коронавирусов и гаммакоронавирусов сохраняются внутриклеточно. В присутствии белка M, трафик спайкового белка изменяется и вместо этого сохраняется в ERGIC, сайте, в котором происходит сборка вируса. В SARS-CoV-2 как белок M, так и белок E модулируют трафик спайкового белка с помощью разных механизмов.
Спайковый белок не требуется для сборки вируса или образования вирусоподобных частиц ; однако наличие шипа может повлиять на размер конверта. Включение белка-шипа в вирионы во время сборки и отпочкования зависит от белок-белковых взаимодействий с белком М через С-концевой хвост. Исследование вирионов с помощью криоэлектронной микроскопии позволяет предположить, что на вирион приходится от 25 до 100 тримеров шипов.
Белок-спайк отвечает за проникновение вируса в клетку-хозяин, что является необходимым ранним этапом репликации вируса. Это важно для репликации. Он выполняет эту функцию в два этапа: сначала связывается с рецептором на поверхности клетки-хозяина посредством взаимодействия с областью S1, а затем сливается вирусная и клеточная мембраны посредством действия области S2. Местоположение слияния варьируется в зависимости от конкретного коронавируса, при этом некоторые из них могут проникать через плазматическую мембрану, а другие - через эндосомы после эндоцитоза.
Взаимодействие рецептор-связывающего домена в области S1 с его рецептором-мишенью на поверхности клетки запускает процесс проникновения вируса. Различные коронавирусы нацелены на разные рецепторы клеточной поверхности, иногда используя молекулы сахара, такие как сиаловые кислоты, или формируя белок-белковые взаимодействия с белками, выставленными на поверхности клетки. Различные коронавирусы сильно различаются по рецепторам-мишеням. Присутствие рецептора-мишени, с которым может связываться S1, является определяющим фактором диапазона хозяев и клеточного тропизма.
Протеолитическое расщепление белка-шипа, иногда известное как «праймирование», необходимо для слияния мембран. По сравнению с другими слитыми белками класса I, этот процесс сложен и требует двух расщеплений в разных сайтах, одно на границе S1 / S2 и одно на сайте S2 'для высвобождения слитого пептида. Коронавирусы различаются, в какой части жизненного цикла вируса происходит это расщепление, особенно расщепление S1 / S2. Многие коронавирусы расщепляются по S1 / S2 перед выходом вируса из вирус-продуцирующей клетки фурином и другими конвертазами пропротеина ; в SARS-CoV-2 в этой позиции присутствует многоосновный сайт расщепления фурином. Другие могут расщепляться внеклеточными протеазами, такими как эластаза, протеазами, находящимися на поверхности клетки после связывания рецептора, или протеазами, обнаруженными в лизосомах после эндоцитоза. Конкретные протеазы, ответственные за это расщепление, зависят от вируса, типа клеток и местной среды. В SARS-CoV, то серин - протеазы TMPRSS2 имеет важное значение для этого процесса, с дополнительным вкладом от цистеинпротеаз катепсина B и катепсина L в эндосомы. Трипсин и трипсин-подобных протеаз также сообщалось внести свой вклад. В SARS-CoV-2 TMPRSS2 является первичной протеазой для расщепления S2 ', и его присутствие, как сообщается, важно для вирусной инфекции.
 Сравнение пре-слияния (оранжевый, голубой) и пост-слияния (красный, темно-синий) конформаций тримерного белка шипа SARS-CoV. В конформации до слияния центральная спираль (оранжевый) и гептадный повтор 1 (HR1, голубой) загнуты друг на друга в антипараллельной ориентации. В конформации после слияния центральная спираль (красный) и последовательность HR1 (темно-синий) реорганизуются с образованием протяженной тримерной спиральной спирали. Вирусная мембрана находится внизу, а мембрана клетки-хозяина вверху. Показаны только ключевые части подблока S2. Из PDB : 6NB6 (до слияния) и PDB : 6M3W (после слияния).
Сравнение пре-слияния (оранжевый, голубой) и пост-слияния (красный, темно-синий) конформаций тримерного белка шипа SARS-CoV. В конформации до слияния центральная спираль (оранжевый) и гептадный повтор 1 (HR1, голубой) загнуты друг на друга в антипараллельной ориентации. В конформации после слияния центральная спираль (красный) и последовательность HR1 (темно-синий) реорганизуются с образованием протяженной тримерной спиральной спирали. Вирусная мембрана находится внизу, а мембрана клетки-хозяина вверху. Показаны только ключевые части подблока S2. Из PDB : 6NB6 (до слияния) и PDB : 6M3W (после слияния). Как и другие слитые белки класса I, спайковый белок в своей конформации до слияния находится в метастабильном состоянии. Резкое конформационное изменение запускается, чтобы вызвать повторное сворачивание гептадных повторов в области S2 в расширенный пучок из шести спиралей, заставляя слитый пептид взаимодействовать с клеточной мембраной и приближая вирусную и клеточную мембраны. Требуется связывание рецептора и протеолитическое расщепление (иногда называемое «прайминг»), но дополнительные триггеры для этого конформационного изменения различаются в зависимости от коронавируса и местной среды. Исследования SARS-CoV in vitro предполагают зависимость от концентрации кальция. В отличие от коронавирусов, вирус инфекционного бронхита, поражающий птиц, может быть вызван одним лишь низким уровнем pH ; для других коронавирусов низкий pH сам по себе не является триггером, но может потребоваться для активности протеаз, которые, в свою очередь, необходимы для слияния. Расположение слияния мембран - на плазматической мембране или в эндосомах - может варьироваться в зависимости от доступности этих триггеров для конформационных изменений. Слияние вирусной и клеточной мембран обеспечивает проникновение генома положительно-смысловой РНК вируса в цитозоль клетки-хозяина, после чего начинается экспрессия вирусных белков.
Помимо слияния мембран вируса и клетки-хозяина, некоторые белки-шипы коронавируса могут инициировать слияние мембран между инфицированными клетками и соседними клетками, образуя синцитии. Такое поведение можно наблюдать в инфицированных клетках в клеточной культуре. Синцитии наблюдались в образцах тканей пациентов, инфицированных SARS-CoV, MERS-CoV и SARS-CoV-2, хотя в некоторых отчетах подчеркивается разница в образовании синцитий между пиками SARS-CoV и SARS-CoV-2, связанная с последовательностью различия вблизи сайта расщепления S1 / S2.
Поскольку он находится на поверхности вируса, спайковый белок является основным антигеном, к которому вырабатываются нейтрализующие антитела. Его обширное гликозилирование может служить гликановым щитом, скрывающим эпитопы от иммунной системы. В связи со вспышкой SARS и пандемией COVID-19 были тщательно изучены антитела к белкам SARS-CoV и SARS-CoV-2. Были идентифицированы антитела к белкам-шипам SARS-CoV и SARS-CoV-2, которые нацелены на эпитопы на рецептор-связывающем домене или мешают процессу конформационных изменений. Большинство антител от инфицированных людей нацелены на рецептор-связывающий домен.
В ответ на COVID-19 пандемии, ряд COVID-19 вакцин были разработаны с использованием различных технологий, в том числе вакцин мРНК и вирусных векторных вакцин. Большинство разработок вакцин нацелено на спайковый белок. Основываясь на методах, ранее использовавшихся в исследованиях вакцин, направленных на респираторно-синцитиальный вирус и SARS-CoV, во многих усилиях по разработке вакцин против SARS-CoV-2 использовались конструкции, которые включают мутации для стабилизации конформации до слияния шипованного белка, способствуя развитию антител против экспонированных эпитопов. в этом подтверждении.
 Казиривимаб (синий) и имдевимаб (оранжевый), взаимодействующие с рецептор-связывающим доменом белка-шипа (розовый).
Казиривимаб (синий) и имдевимаб (оранжевый), взаимодействующие с рецептор-связывающим доменом белка-шипа (розовый). Моноклональные антитела, нацеленные на спайковый белок, были разработаны для лечения COVID-19. По состоянию на 8 июля 2021 г. три продукта с моноклональными антителами получили разрешение на экстренное использование в США: бамланивимаб / этесевимаб, казиривимаб / имдевимаб и сотровимаб. Бамланивимаб / этесевимаб не рекомендован в США из-за увеличения числа вариантов SARS-CoV-2, которые менее чувствительны к этим антителам.
На протяжении COVID-19 пандемии, то геном ТОРС-коронавирус-2 вирусов был секвенирован много раз, в результате идентификации тысяч различных вариантов. Многие из них обладают мутациями, которые изменяют аминокислотную последовательность белка-шипа. Согласно анализу Всемирной организации здравоохранения, проведенному в июле 2020 года, ген spike ( S) был вторым по частоте мутацией в геноме после ORF1ab (который кодирует большинство неструктурных белков вируса). Скорость эволюции гена шипа выше, чем наблюдаемая в геноме в целом. Анализ геномов SARS-CoV-2 позволяет предположить, что некоторые участки в последовательности белка-шипа, особенно в рецептор-связывающем домене, имеют эволюционное значение и подвергаются положительному отбору.
Спайковые мутации белков вызывают озабоченность, поскольку они могут влиять на инфекционность или трансмиссивность или способствовать ускользанию от иммунитета. Мутация D 614 G возникла независимо во множестве вирусных линий и стала доминирующей среди секвенированных геномов; он может иметь преимущества в отношении инфекционности и трансмиссивности, возможно, из-за увеличения плотности шипов на вирусной поверхности, увеличения доли компетентных к связыванию конформаций или повышения стабильности. Мутации в положении E 484, особенно E 484 K, были связаны с иммунным ускользанием и снижением связывания антител.
Во время COVID-19 пандемии, против вакцинации дезинформации о COVID-19, распространенном на социальных медиа - платформ, связанных с ролью шипа белка в COVID-19 вакцин. Говорят, что спайковые белки опасно « цитотоксичны », а содержащие их мРНК вакцины, следовательно, опасны сами по себе. Спайковые белки не цитотоксичны и не опасны. Сообщалось также, что спайковые белки «сбрасываются» вакцинированными людьми, что является ошибочной ссылкой на феномен индуцированного вакциной выделения вируса, который является редким эффектом живых вирусных вакцин, в отличие от вакцин, используемых для COVID-19. «Отбрасывание» белков-шипов невозможно.
В слитые белки класса I, группа, у которой хорошо охарактеризованных примеры включают белок коронавируса шипа, вируса гриппа гемагглютинин и ВИЧ gp41, как полагают, эволюционно связаны. Участок S2 белка-шипа, ответственного за слияние мембран, более консервативен, чем участок S1, ответственный за взаимодействия с рецепторами. Похоже, что регион S1 подвергся значительному диверсификационному отбору.
В области S1 N-концевой домен более консервативен, чем C-концевой домен. НТД в галектин -подобных белков складка предполагает связь с структурно подобных клеточных белков, из которых, возможно, развились через захвата генов от хозяина. Было высказано предположение, что CTD мог развиться из NTD в результате дупликации генов. Открытая поверхность CTD, уязвимая для иммунной системы хозяина, может подвергнуть эту область высокому селективному давлению. Сравнение структур различных CTD коронавирусов предполагает, что они могут подвергаться диверсификации, и в некоторых случаях отдаленно родственные коронавирусы, использующие один и тот же рецептор на клеточной поверхности, могут делать это в результате конвергентной эволюции.