
Войти
 Цикл Кори
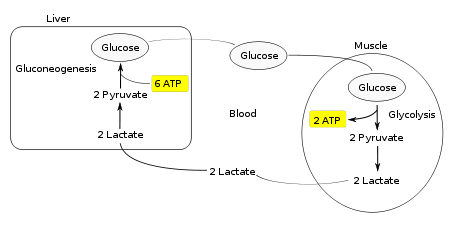
Цикл Кори Цикл Кори (также известный как цикл молочной кислоты ), названный в честь его первооткрывателей, Карла Фердинанда Кори и Герти Кори, представляет собой метаболический путь, в котором лактат вырабатывается анаэробным гликолизом в мышцах транспортируется в печень и превращается в глюкозу, которая затем возвращается в мышцы и циклически метаболизируется обратно в лактат.
 Карл Кори и Герти Кори совместно получили Нобелевскую премию 1947 года по физиологии и медицине, а не за открытие Цикл Кори, но для их открытия процесса каталитического превращения гликогена.
Карл Кори и Герти Кори совместно получили Нобелевскую премию 1947 года по физиологии и медицине, а не за открытие Цикл Кори, но для их открытия процесса каталитического превращения гликогена. Мышечная активность требует АТФ, который обеспечивается расщеплением гликогена в скелете мышцы. Распад гликогена, известный как гликогенолиз, высвобождает глюкозу в форме глюкозо-1-фосфата (G1P). G1P превращается в G6P с помощью фосфоглюкомутазы. G6P легко вводится в гликолиз (или может переходить в пентозофосфатный путь, если концентрация G6P высока) процесс, который обеспечивает АТФ мышечными клетками в виде источник энергии. Во время мышечной активности необходимо постоянно пополнять запасы АТФ. Когда подача кислорода достаточна, эта энергия поступает от подачи пирувата, одного продукта гликолиза, в цикл лимонной кислоты.
. Когда подача кислорода недостаточна, обычно во время интенсивной мышечной активности энергия должна высвобождаться посредством анаэробного метаболизма. Молочнокислая ферментация превращает пируват в лактат с помощью лактатдегидрогеназы. Что наиболее важно, ферментация регенерирует NAD, поддерживая его концентрацию, поэтому могут происходить дополнительные реакции гликолиза. На стадии ферментации НАДН, полученный в результате гликолиза, окисляется обратно до НАД, передавая два электрона от НАДН для восстановления пирувата до лактата. (За подробностями обращайтесь к основным статьям по гликолизу и ферментации.)
Вместо того, чтобы накапливаться внутри мышечных клеток, лактат, произведенный анаэробной ферментацией, поглощается печень. Это инициирует вторую половину цикла Кори. В печени происходит глюконеогенез. С интуитивной точки зрения глюконеогенез обращает вспять как гликолиз, так и ферментацию, превращая лактат сначала в пируват, а затем обратно в глюкозу. Затем глюкоза поступает в мышцы через кровоток ; он готов к дальнейшим реакциям гликолиза. Если мышечная активность прекратилась, глюкоза используется для пополнения запасов гликогена посредством гликогенеза.
В целом на этапах гликолиза цикла вырабатываются 2 молекулы АТФ при затратах 6 молекул АТФ, потребляемых на этапах глюконеогенеза. Каждая итерация цикла должна поддерживаться чистым потреблением 4 молекул АТФ. В результате цикл не может продолжаться бесконечно. Интенсивное потребление молекул АТФ в цикле Кори сдвигает метаболическую нагрузку с мышц на печень.
Важность цикла основана на предотвращении лактоацидоза в мышцах в анаэробных условиях. Однако обычно, прежде чем это произойдет, молочная кислота перемещается из мышц в печень.
Цикл также важен для производства АТФ, источника энергии, во время мышечной активности. Цикл Кори функционирует более эффективно, когда мышечная активность прекращается. Это позволяет погашать кислородный долг, так что цикл лимонной кислоты и цепь переноса электронов могут производить энергию с максимальной эффективностью.
Цикл Кори является гораздо более важным источником субстрата для глюконеогенеза чем еда. Вклад лактата цикла Кори в общую продукцию глюкозы увеличивается с периодом голодания до выхода на плато. В частности, через 12, 20 и 40 часов голодания добровольцев-людей на глюконеогенез приходится 41%, 71% и 92% продукции глюкозы, но вклад лактата цикла Кори в глюконеогенез составляет 18%, 35% и 36%. % соответственно. Оставшееся производство глюкозы происходит за счет распада белка, мышечного гликогена и глицерина в результате липолиза.
Препарат метформин может вызывать лактоацидоз у пациентов с почечной недостаточностью потому что метформин ингибирует печеночный глюконеогенез цикла Кори, особенно комплекс 1 дыхательной цепи митохондрий. Накопление лактата и его субстратов для производства лактата, пирувата и аланина, приводит к избытку лактата. Обычно избыток лактата выводится почками, но у пациентов с почечной недостаточностью почки не могут справиться с избытком молочной кислоты.