
Войти
 Эта дупликация гена создала вариацию количества копий. Хромосома теперь имеет две копии этого участка ДНК, а не одну.
Эта дупликация гена создала вариацию количества копий. Хромосома теперь имеет две копии этого участка ДНК, а не одну. Вариация количества копий (CNV ) - это явление, при котором участки генома повторяются и количество повторов в геноме варьируется от человека к человеку. Вариация числа копий - это тип структурной вариации : в частности, это тип события дупликации или удаления, который влияет на значительное количество пар оснований. Примерно две трети всего генома человека могут состоять из повторов, а 4,8–9,5% генома человека можно классифицировать как вариации числа копий. У млекопитающих вариации количества копий играют важную роль в создании необходимых вариаций в популяции, а также фенотипа заболевания.
Вариации количества копий в целом можно разделить на две основные группы: короткие повторы и длинные повторы. Однако четких границ между двумя группами нет, и классификация зависит от природы интересующих локусов. Короткие повторы включают в основном двухнуклеотидные повторы (два повторяющихся нуклеотида, например, A-C-A-C-A-C...) и трехнуклеотидные повторы. Длинные повторы включают повторы целых генов. Эта классификация, основанная на размере повтора, является наиболее очевидным типом классификации, поскольку размер является важным фактором при изучении типов механизмов, которые с наибольшей вероятностью вызывают повторы, следовательно, вероятное влияние этих повторов на фенотип.
Одним из наиболее известных примеров вариации короткого количества копий является трехнуклеотидный повтор пар оснований CAG в гене хантингтина (не в гене Хантингтона), гене, который отвечает за неврологическое расстройство болезнь Хантингтона. В этом конкретном случае, если тринуклеотид CAG повторится более 36 раз, болезнь Хантингтона, вероятно, разовьется у человека и, вероятно, будет унаследована его или ее потомством. Число повторов тринуклеотида CAG коррелирует с возрастом начала болезни Хантингтона. Эти типы коротких повторов часто считаются следствием ошибок в активности полимеразы во время репликации, включая проскальзывание полимеразы, переключение шаблона и переключение вилки, которые будут подробно обсуждаться позже. Короткий размер повтора в этих вариациях числа копий приводит к ошибкам в полимеразе, поскольку эти повторяющиеся области склонны к неправильному распознаванию полимеразой, и реплицированные области могут реплицироваться снова, что приводит к дополнительным копиям повтора. Кроме того, если эти тринуклеотидные повторы находятся в одной и той же рамке считывания в кодирующей части гена, это может привести к длинной цепи той же аминокислоты., возможно, создавая белковые агрегаты в клетке, и если эти короткие повторы попадают в некодирующую часть гена, это может повлиять на экспрессию гена и регуляцию. С другой стороны, в геноме реже выявляется переменное количество повторов целых генов. Одним из примеров повтора целого гена является ген альфа-амилазы 1 (AMY1), который кодирует альфа-амилазу, которая имеет значительные различия в количестве копий между разными популяциями с разными диетами. Хотя конкретный механизм, который позволяет гену AMY1 увеличивать или уменьшать количество его копий, все еще является предметом обсуждения, некоторые гипотезы предполагают, что негомологичное соединение концов или опосредованное микрогомологией соединение концов, вероятно, отвечает за повторы целого гена. Повторение целых генов немедленно влияет на экспрессию этого конкретного гена, и тот факт, что изменение числа копий гена AMY1 связано с диетой, является замечательным примером недавней эволюционной адаптации человека. Хотя это общие группы, в которые сгруппированы вариации количества копий, точное количество вариаций количества копий пар оснований, на которые влияют, зависит от конкретных интересующих локусов. В настоящее время, используя данные всех зарегистрированных вариаций количества копий, средний размер варианта количества копий составляет около 118 КБ, а медиана - около 18 КБ.
С точки зрения структурной архитектуры вариаций количества копий, исследования предложили и определены горячие точки в геноме, где вариации числа копий в четыре раза больше. Эти горячие точки были определены как области, содержащие длинные повторы, которые на 90–100% похожи, известные как сегментарные дупликации либо тандем, либо перемежающиеся, и, что наиболее важно, эти горячие точки имеют повышенную скорость хромосомная перестройка. Считалось, что эти крупномасштабные хромосомные перестройки вызывают нормальные вариации и генетические заболевания, включая вариации числа копий. Более того, эти горячие точки вариации числа копий согласованы во многих популяциях с разных континентов, подразумевая, что эти горячие точки были либо независимо приобретены всеми популяциями и передавались из поколения в поколение, либо они были приобретены на ранней стадии эволюции человека до того, как популяции разделились, последнее кажется более вероятно. И, наконец, в геноме, по-видимому, не происходит пространственных искажений местоположения, в котором вариации числа копий наиболее плотно распределены. Хотя изначально с помощью флуоресцентной гибридизации in situ и микросателлитного анализа было обнаружено, что повторы числа копий локализованы в областях, которые часто повторяются, таких как теломеры, центромеры и гетерохроматин, недавние полногеномные исследования пришли к другому выводу. А именно, в субтеломерных областях и перицентромерных областях обнаруживается большинство горячих точек хромосомной перестройки, и не наблюдается значительного увеличения вариаций числа копий в этой области. Кроме того, эти области горячих точек хромосомной перестройки не имеют уменьшенного количества генов, что опять же означает минимальное пространственное смещение геномного местоположения вариаций количества копий.
Число копий Изначально считалось, что вариации занимают чрезвычайно малую и незначительную часть генома посредством цитогенетических наблюдений. Вариации количества копий обычно были связаны только с небольшими тандемными повторами или конкретными генетическими нарушениями, поэтому вариации количества копий первоначально исследовались только с точки зрения конкретных локусов. Однако технологические разработки привели к появлению все большего числа высокоточных способов определения и изучения вариаций количества копий. Вариации числа копий первоначально изучались цитогенетическими методами, которые позволяют наблюдать физическую структуру хромосомы. Одним из таких методов является флуоресцентная гибридизация in situ (FISH), которая включает в себя введение флуоресцентных зондов, требующих высокой степени комплементарности в геном для связывания. Сравнительная геномная гибридизация также обычно использовался для обнаружения вариаций числа копий с помощью визуализации флуорофора и последующего сравнения длины хромосом. Одним из основных недостатков этих ранних методов является то, что разрешение генома относительно низкое, и могут быть обнаружены только большие повторы, такие как повторы целого гена.
Недавние достижения в технологиях геномики привели к появлению многих важных методов, которые имеют чрезвычайно высокое геномное разрешение, и, как результат, сообщалось о растущем количестве вариаций числа копий в геноме. Первоначально эти достижения включали использование массива бактериальной искусственной хромосомы (ВАС) с интервалом около 1 мегабазы по всему гену, ВАС также могут обнаруживать вариации числа копий в горячих точках реаранжировки, что позволяет обнаруживать 119 новых вариаций числа копий. высокопроизводительное геномное секвенирование произвело революцию в области геномики человека, и были выполнены in silico исследования для обнаружения вариаций числа копий в геноме. Контрольные последовательности сравнивали с другими представляющими интерес последовательностями с использованием фосмид, строго контролируя, чтобы клоны фосмид имели размер 40 т.п.н. Считывание конца секвенирования предоставит адекватную информацию для выравнивания эталонной последовательности с интересующей последовательностью, и любые несовпадения легко заметны, что позволяет сделать вывод, что это вариации числа копий в этой области клона. Этот тип метода обнаружения обеспечивает высокое геномное разрешение и точное расположение повтора в геноме, а также может обнаруживать другие типы структурных вариаций, такие как инверсии.
Кроме того, существует другой способ обнаружения вариации числа копий использует однонуклеотидный полиморфизм (SNP). Из-за обилия человеческих данных SNP направление обнаружения вариации количества копий изменилось, чтобы использовать эти SNP. Опираясь на тот факт, что рекомбинация человека является относительно редкой и что многие события рекомбинации происходят в определенных областях генома, известных как горячие точки рекомбинации, неравновесие по сцеплению можно использовать для идентификации вариаций количества копий. Были предприняты попытки связать вариации числа копий с конкретными SNP гаплотипа путем анализа неравновесия по сцеплению, используя эти ассоциации, можно распознать вариации числа копий в геноме, используя SNP в качестве маркеров. Секвенирование следующего поколения В настоящее время все чаще используются методы секвенирования с коротким и длинным считыванием, которые начали заменять методы на основе массивов для обнаружения вариаций количества копий. В отличие от методов на основе массивов, методы обнаружения на основе секвенирования легко идентифицируют другие классы структурной вариации, такие как инверсии и транслокации.
Существует два основных типа молекулярных механизмов образования вариаций числа копий: на основе гомолога и на основе негомолога. Хотя было выдвинуто много предположений, большинство этих теорий являются предположениями и предположениями. Нет убедительных доказательств того, что конкретное изменение количества копий коррелирует с конкретным механизмом.
 Схематическое изображение неаллельной гомологичной рекомбинации. Здесь Ген X представляет интересующий ген, а черная линия представляет хромосому. Когда две гомологичные хромосомы неправильно выровнены и происходит рекомбинация, это может привести к дублированию гена.
Схематическое изображение неаллельной гомологичной рекомбинации. Здесь Ген X представляет интересующий ген, а черная линия представляет хромосому. Когда две гомологичные хромосомы неправильно выровнены и происходит рекомбинация, это может привести к дублированию гена. Одна из наиболее известных теорий, которая приводит к вариациям числа копий, а также к делециям и инверсиям, является неаллельные гомологичные рекомбинации. Во время мейотической рекомбинации гомологичные хромосомы образуют пары и образуют двухконцевые двухцепочечные разрывы, приводящие к соединениям Холлидея. Однако в аберрантном механизме во время образования соединений Холлидея двухцепочечные разрывы смещены, и кроссовер попадает в неаллельные положения на одной и той же хромосоме. Когда соединение Холлидея разрешено, событие неравного кроссинговера позволяет переносить генетический материал между двумя гомологичными хромосомами, и в результате часть ДНК на обоих гомологах повторяется. Поскольку повторяющиеся области больше не сегрегируют независимо, дублированная область хромосомы наследуется. Другой тип механизма, основанного на гомологичной рекомбинации, который может приводить к изменению числа копий, известен как репликация, индуцированная разрывом. Когда в геноме неожиданно возникает двухцепочечный разрыв, клетка активирует пути, которые опосредуют восстановление разрыва. Ошибки при восстановлении разрыва, аналогичные неаллельной гомологичной рекомбинации, могут приводить к увеличению числа копий конкретной области генома. Во время восстановления двухцепочечного разрыва сломанный конец может вторгнуться в его гомологичную хромосому вместо того, чтобы присоединиться к исходной цепи. Как и в механизме неаллельной гомологичной рекомбинации, дополнительная копия конкретной области переносится на другую хромосому, что приводит к событию дупликации. Кроме того, обнаружено, что белки когезин помогают в системе репарации двухцепочечных разрывов за счет зажима двух концов в непосредственной близости, что предотвращает межхромосомную инвазию концов. Если по какой-либо причине, такой как активация рибосомной РНК, активность когезина нарушается, тогда может наблюдаться локальное увеличение ошибок восстановления двухцепочечных разрывов.
Другой класс возможных механизмов, которые предполагаются приводить к вариациям количества копий не является гомологичным. Чтобы различать этот механизм и механизмы, основанные на гомологии, необходимо понимать концепцию гомологии. Гомологичное спаривание хромосом происходит с использованием нитей ДНК, которые очень похожи друг на друга (~ 97%), и эти цепи должны быть длиннее определенной длины, чтобы избежать коротких, но очень похожих пар. С другой стороны, негомологичные пары основываются только на нескольких парах оснований сходства между двумя цепями, поэтому возможен обмен или дублирование генетических материалов в процессе восстановления двухцепочечных цепей на негомологичной основе.
Одним из типов механизма на негомологичной основе является механизм негомологичного соединения концов или микрогомологичного соединения концов. Эти механизмы также участвуют в репарации двухцепочечных разрывов, но не требуют гомологии или ограниченной микрогомологии. Когда эти цепи восстанавливаются, часто в восстановленную цепь добавляются небольшие делеции или вставки. Возможно, что ретротранспозоны вставляются в геном через эту систему репарации. Если ретротранспозоны вставлены в неаллельную позицию на хромосоме, мейотическая рекомбинация может привести к рекомбинации вставки в ту же цепь, что и уже существующая копия той же области. Другим механизмом является цикл разрыва-слияния-мостика, который включает сестринские хроматиды, которые обе потеряли свою теломерную область из-за двухцепочечных разрывов. Предполагается, что эти сестринские хроматиды будут сливаться вместе с образованием одной дицентрической хромосомы, а затем разделяться на два разных ядра. Поскольку разделение дицентрической хромосомы вызывает двухцепочечный разрыв, концевые области могут сливаться с другими двухцепочечными разрывами и повторять цикл. Слияние двух сестринских хроматид может вызвать инвертированную дупликацию, и когда эти события повторяются в течение всего цикла, инвертированная область будет повторяться, что приведет к увеличению числа копий. Последний механизм, который может привести к изменению числа копий, - это проскальзывание полимеразы, которое также известно как переключение шаблона. Во время нормальной репликации ДНК полимераза на отстающей цепи необходима для непрерывного разжимания и повторного зажима области репликации. Когда мелкомасштабные повторы в последовательности ДНК уже существуют, полимераза может быть «сбита с толку», когда она повторно зажимается для продолжения репликации, и вместо того, чтобы зажимать правильные пары оснований, она может сдвинуть несколько пар оснований и реплицировать часть повторяющейся регион снова. Обратите внимание, что хотя это было экспериментально обнаружено и является широко принятым механизмом, молекулярные взаимодействия, которые привели к этой ошибке, остаются неизвестными. Кроме того, поскольку этот тип механизма требует, чтобы полимераза прыгала вокруг нити ДНК, и маловероятно, что полимераза сможет повторно зажимать другой локус на расстоянии нескольких тысяч пар оснований, поэтому это более применимо к коротким повторам, таким как бинуклеотидные или три -нуклеотидные повторы.
 Хронология изменения рациона гомининов в периоды позднего палеолита, мезолита и неолита. Как видно, корнеплоды, богатые крахмалом, потреблялись около 20000 лет назад, когда количество диплоидных генов AMY1 увеличивалось.
Хронология изменения рациона гомининов в периоды позднего палеолита, мезолита и неолита. Как видно, корнеплоды, богатые крахмалом, потреблялись около 20000 лет назад, когда количество диплоидных генов AMY1 увеличивалось. Амилаза - это фермент в слюне, который отвечает за расщепление крахмал в моносахариды, и один тип амилазы кодируется геном альфа-амилазы (AMY1). Локус AMY1, как и фермент амилаза, является одним из наиболее широко изученных и секвенированных генов в геноме человека. Его гомологи также обнаружены у других приматов, и поэтому вполне вероятно, что ген приматов AMY1 является предком гена AMY1 человека и был адаптирован на ранних этапах эволюции приматов. AMY1 - один из наиболее хорошо изученных генов, который имеет широкий диапазон переменного числа копий в разных популяциях человека. Ген AMY1 также является одним из немногих изученных генов, которые продемонстрировали убедительные доказательства корреляции его функции белка с числом его копий. Число копий, как известно, изменяет уровни транскрипции, а также трансляции конкретного гена, однако исследования показали, что взаимосвязь между уровнями белка и количеством копий варьируется. В генах AMY1 американцев европейского происхождения обнаружено, что концентрация амилазы в слюне тесно коррелирует с числом копий гена AMY1. В результате была выдвинута гипотеза, что количество копий гена AMY1 тесно коррелирует с его функцией белка, которая заключается в переваривании крахмала.
Было обнаружено, что количество копий гена AMY1 коррелирует с различными уровнями крахмал в рационах разных популяций. 8 Популяции с разных континентов были разделены на рационы с высоким содержанием крахмала и рационы с низким содержанием крахмала, и их количество копий гена AMY1 было визуализировано с использованием FISH с высоким разрешением и qPCR. Было обнаружено, что популяции с высоким содержанием крахмала, которые состоят из популяций японцев, хадза и европейцев Америки, имели значительно более высокое (в 2 раза выше) среднее число копий AMY1, чем популяции с низким содержанием крахмала, включая популяции биака, мбути, датог, якуты. Была высказана гипотеза, что уровень крахмала в обычном рационе, субстрате для AMY1, может напрямую влиять на количество копий гена AMY1. Поскольку был сделан вывод, что количество копий AMY1 напрямую коррелирует с амилазой слюны, чем больше крахмала присутствует в ежедневном рационе населения, тем более благоприятным с точки зрения эволюции является наличие нескольких копий гена AMY1. Ген AMY1 был первым геном, предоставившим убедительные доказательства эволюции на уровне молекулярной генетики. Более того, с помощью сравнительной геномной гибридизации вариации числа копий для всех геномов японской популяции сравнивали с таковой у якутской популяции. Было обнаружено, что вариация количества копий гена AMY1 значительно отличается от вариации количества копий в других генах или областях генома, что позволяет предположить, что ген AMY1 находился под сильным селективным давлением, которое практически не влияло на другую копию. количество вариаций. Наконец, вариабельность длины 783 микросателлитов между двумя популяциями сравнивали с вариабельностью числа копий гена AMY1. Было обнаружено, что диапазон числа копий гена AMY1 был больше, чем у более 97% исследованных микросателлитов. Это означает, что естественный отбор сыграл значительную роль в формировании среднего количества генов AMY1 в этих двух популяциях. Однако, поскольку были изучены только 6 популяций, важно учитывать возможность того, что в их рационе или культуре могут быть другие факторы, помимо крахмала, которые влияли на количество копий AMY1.
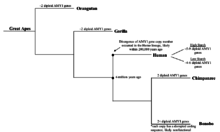 Упрощенное филогенетическое дерево происхождения великих обезьян и количество диплоидных генов AMY1, которые есть у каждого вида. Показано, что количество гена AMY1 увеличивается после разделения с линией шимпанзе.
Упрощенное филогенетическое дерево происхождения великих обезьян и количество диплоидных генов AMY1, которые есть у каждого вида. Показано, что количество гена AMY1 увеличивается после разделения с линией шимпанзе. Хотя неясно, когда стало увеличиваться количество копий гена AMY1, известно и подтверждено, что ген AMY1 существовал у ранних приматов. У шимпанзе, ближайших эволюционных родственников человека, было обнаружено 2 диплоидных копии гена AMY1, идентичной по длине гену AMY1 человека, что значительно меньше, чем у человека. люди. С другой стороны, бонобо, также близкий родственник современного человека, обнаружил более 2 диплоидных копий гена AMY1. Тем не менее, гены AMY1 бонобо были секвенированы и проанализированы, и было обнаружено, что кодирующие последовательности генов AMY1 нарушаются, что может привести к продукции дисфункциональной амилазы слюны. Из результатов можно сделать вывод, что увеличение числа копий AMY1 бонобо, вероятно, не коррелирует с количеством крахмала в их рационе. Кроме того, была высказана гипотеза, что увеличение числа копий началось недавно, во время ранней эволюции гоминина, поскольку ни у одного из человекообразных обезьян не было более двух копий гена AMY1, продуцирующего функциональный белок. Кроме того, было высказано предположение, что увеличение количества копий AMY1 началось около 20000 лет назад, когда люди перешли от образа жизни охотника-собирателя к сельскохозяйственному обществу, что также было тогда, когда люди полагались на много корнеплодов с высоким содержанием крахмала. Эта гипотеза, хотя и логична, не имеет экспериментальных доказательств из-за трудностей со сбором информации об изменении рациона человека, особенно о корнеплодах с высоким содержанием крахмала, поскольку их нельзя непосредственно наблюдать или тестировать. Недавние открытия в области секвенирования ДНК позволили исследователям с определенной степенью точности секвенировать более старую ДНК, такую как ДНК неандертальцев. Возможно, секвенирование ДНК неандертальцев может предоставить временной маркер увеличения числа копий гена AMY1 и дать представление о рационе человека и эволюции генов.
В настоящее время неизвестно, какой механизм привел к начальной дупликации гена амилазы, и это может означать, что вставка последовательностей ретровирусов была вызвана негомологичным соединением концов, что вызвали дупликацию гена AMY1. Однако в настоящее время нет доказательств в поддержку этой теории, и поэтому эта гипотеза остается предположением. Недавнее происхождение мультикопийного гена AMY1 подразумевает, что в зависимости от окружающей среды количество копий гена AMY1 может увеличиваться и уменьшаться очень быстро по сравнению с генами, которые не взаимодействуют напрямую с окружающей средой. Ген AMY1 является прекрасным примером того, как дозировка гена влияет на выживаемость организма в данной среде. Множественные копии гена AMY1 дают тем, кто в большей степени полагается на диету с высоким содержанием крахмала, эволюционное преимущество, поэтому в популяции сохраняется высокое число копий гена.
Среди нейронов в головном мозге человека, часто возникают соматически полученные вариации числа копий. Вариации числа копий демонстрируют широкую изменчивость (от 9 до 100% нейронов мозга в разных исследованиях). Большинство изменений имеют размер от 2 до 10 МБ, при этом количество удалений намного превышает количество амплификаций. Вариации числа копий в клетках мозга выше, чем в клетках других типов. Вероятным источником вариации числа копий является неправильная репарация повреждений ДНК..
Геномная дупликация и тройная репликация гена, по-видимому, являются редкой причиной болезни Паркинсона, хотя и более распространены, чем точечные мутации.
 Возможный механизм того, как множественные копии гена могут привести к созданию семейства белков в течение многих лет при естественном отборе. Здесь ген X представляет собой представляющий интерес ген, который дуплицирован, а ген X1 и ген X2 - это гены, которые приобрели мутации и стали функционально отличными от гена X.
Возможный механизм того, как множественные копии гена могут привести к созданию семейства белков в течение многих лет при естественном отборе. Здесь ген X представляет собой представляющий интерес ген, который дуплицирован, а ген X1 и ген X2 - это гены, которые приобрели мутации и стали функционально отличными от гена X. Недавно обсуждалась связь вариаций числа копий с семействами генов.. Семейства генов определяются как набор родственных генов, которые выполняют сходные функции, но имеют незначительные временные или пространственные различия, и эти гены, вероятно, происходят от одного предкового гена. Основная причина, по которой вариации числа копий связаны с семействами генов, заключается в том, что существует вероятность того, что гены в семье могли происходить от одного предкового гена, который дублировался в разные копии. Мутации со временем накапливаются в генах, и с естественным отбором, воздействующим на гены, некоторые мутации приводят к преимуществам окружающей среды, позволяя этим генам наследоваться, и в конечном итоге чистые генные семейства отделяются. Примером семейства генов, которое могло быть создано из-за вариаций числа копий, является семейство генов глобина. Семейство глобиновых генов представляет собой сложную сеть генов, состоящую из альфа и бета генов глобина, включая гены, которые экспрессируются как у эмбрионов, так и у взрослых, а также псевдогенов. Все эти глобиновые гены в семействе глобинов хорошо законсервированы и отличаются только небольшой частью гена, что указывает на то, что они произошли от общего предкового гена, возможно, из-за дупликации исходного гена глобина.
Исследования показал, что вариации числа копий значительно чаще встречаются в генах, кодирующих белки, которые непосредственно взаимодействуют с окружающей средой, чем белки, которые участвуют в основных клеточных действиях. Было высказано предположение, что эффект дозировки гена, сопровождающий изменение числа копий, может приводить к пагубным эффектам, если нарушаются основные клеточные функции, поэтому белки, участвующие в клеточных путях, подвергаются сильной очищающей селекции. Кроме того, белки функционируют вместе и взаимодействуют с белками других путей, поэтому важно рассматривать влияние естественного отбора на биомолекулярные пути, а не на отдельные белки. При этом было обнаружено, что белки на периферии пути обогащены вариациями числа копий, тогда как белки в центре пути обеднены вариациями числа копий. Было объяснено, что белки на периферии пути взаимодействуют с меньшим количеством белков, и поэтому изменение дозировки белка, вызванное изменением числа копий, может иметь меньшее влияние на общий результат клеточного пути.
В В последние несколько лет исследователи, похоже, сместили свое внимание с обнаружения, локализации и секвенирования вариаций числа копий на углубленный анализ роли этих вариаций числа копий в геноме человека и в природе в целом. Доказательства необходимы для дальнейшего подтверждения взаимосвязи между вариациями числа копий и семействами генов, а также той роли, которую естественный отбор играет в формировании этих отношений и изменений. Кроме того, исследователи также стремятся выяснить молекулярные механизмы, участвующие в вариациях числа копий, поскольку это может дать важную информацию о структурных вариациях в целом. Сделав шаг назад, можно сказать, что область структурных вариаций в геноме человека становится быстро развивающейся темой исследований. Эти данные исследований не только могут предоставить дополнительные доказательства эволюции и естественного отбора, но также могут быть использованы для разработки методов лечения широкого спектра генетических заболеваний.