
Войти
| Коннексин | |||||||||
|---|---|---|---|---|---|---|---|---|---|
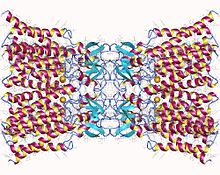 Додекамер Коннексин-26. Щелевое соединение, состоящее из двенадцати идентичных белков коннексина, по шесть в мембране каждой клетки. Каждая из этих шести единиц представляет собой отдельный полипептид, который проходит через мембрану четыре раза (называемые четырехпроходными трансмембранными белками). Додекамер Коннексин-26. Щелевое соединение, состоящее из двенадцати идентичных белков коннексина, по шесть в мембране каждой клетки. Каждая из этих шести единиц представляет собой отдельный полипептид, который проходит через мембрану четыре раза (называемые четырехпроходными трансмембранными белками). | |||||||||
| Идентификаторы | |||||||||
| Символ | Коннексин | ||||||||
| Pfam | PF00029 | ||||||||
| InterPro | IPR013092 | ||||||||
| PROSITE | PDOC00341 | ||||||||
| TCDB | 1.A.24 | ||||||||
| суперсемейство OPM | 194 | ||||||||
| белок OPM | 2zw3 | ||||||||
| |||||||||
коннексины (Cx) (TC # 1.A.24 ) или щелевые соединения белки, являются структурно родственными трансмембранными белками, которые собираются с образованием щелевых соединений позвоночных. Совершенно другое семейство белков, иннексины, образуют щелевые соединения у беспозвоночных. Каждый щелевой стык состоит из двух полуканалов, или коннексонов, которые состоят из гомо- или гетерогексамерных массивов коннексинов, и коннексон в одной плазматической мембране стыкуется встык с коннексоном в мембране близко противостоящая ячейка. Гемиканал состоит из шести субъединиц коннексина, каждая из которых состоит из четырех трансмембранных сегментов. Щелевые соединения необходимы для многих физиологических процессов, таких как скоординированная деполяризация сердечной мышцы, правильное эмбриональное развитие и управляемая реакция микрососудов. По этой причине мутации в генах, кодирующих коннексин, могут приводить к функциональным нарушениям и аномалиям развития.
Коннексины обычно называют в соответствии с их молекулярными массами, например Cx26 представляет собой белок коннексин 26 кДа. Конкурирующей номенклатурой является система белков щелевых соединений, в которой коннексины сортируются по их α (GJA) и β (GJB) формам, а дополнительные коннексины сгруппированы в группы C, D и E, за которыми следуют идентифицирующие число, например GJA1 соответствует Cx43. После голосования на конференции Gap Junction Conference (2007) в Эльсиноре сообщество согласилось использовать систему номенклатуры GJ для генов, кодирующих коннексины, но пожелало сохранить номенклатуру коннексинов для кодируемых белков с использованием веса человеческого белка для нумерации. ортологичных белков.
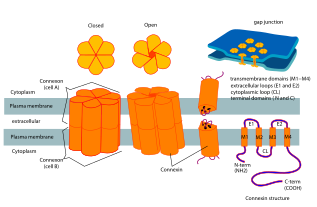
Коннексины содержат четыре высокоупорядоченных трансмембранных сегмента (TMS), в основном неструктурированные цитоплазматические концы C и N, цитоплазматическую петлю (CL) и две внеклеточные петли (EL -1) и (EL-2). Коннексины собираются в группы по шесть, чтобы сформировать полуканалы или коннексоны, а два полуканала затем объединяются, образуя щелевое соединение.
Доступна кристаллическая структура канала щелевого соединения, образованного человеческим Cx26 (также известный как GJB2) с разрешением 3,5 Å. Карта плотности показывала два межмембранных полуканала и расположение четырех TMS шести протомеров, образующих каждый полуканал. Гемиканалы имеют положительно заряженный вход в цитоплазму, воронку, отрицательно заряженный трансмембранный путь и внеклеточную полость. Пора сужается у воронки, которая образована шестью аминоконцевыми спиралями, выстилающими стенку канала, что, таким образом, определяет ограничение размера молекулы на входе в канал.
Семейство генов коннексина разнообразно: двадцать один идентифицированный член в секвенированном геноме человека и двадцать у мыши (девятнадцать из которых являются ортологичными парами). Обычно они весят от 25 до 60 кДа и имеют среднюю длину 380 аминокислот. Было обнаружено, что различные коннексины объединяются как в гомомерные, так и в гетеромерные щелевые переходы, каждый из которых может проявлять разные функциональные свойства, включая проводимость пор, избирательность по размеру, селективность заряда, стробирование напряжения и химическое стробирование.
Замечательным аспектом коннексинов является то, что у них относительно короткий период полураспада, составляющий всего несколько часов. Результатом является наличие динамического цикла, в котором синтезируются и заменяются коннексины. Было высказано предположение, что такая короткая продолжительность жизни позволяет более точно регулировать физиологические процессы, например, в миометрии.
, поскольку они транслируются рибосомами., коннексины вставляются в мембрану эндоплазматического ретикулума (ER). Именно в ER коннексины сворачиваются правильно, образуя две внеклеточные петли, EL-1 и EL-2. Также именно в ER начинается олигомеризация молекул коннексина в полуканалы, процесс, который также может продолжаться в промежуточном компартменте UR-Golgi. Расположение этих гемиканалов может быть гомотипическим, гетеротипическим и комбинированным гетеротипическим / гетеромерным. После выхода из ER и прохождения через ERGIC свернутые коннексины обычно попадают в сеть цис-Гольджи. Однако некоторые коннексины, такие как Сх26, могут транспортироваться независимо от Гольджи.
После вставки в плазматическую мембрану клетки гемиканалы свободно диффундируют в липидном бислое.. С помощью специфических белков, в основном кадгеринов, полуканалы могут стыковаться с полуканалами соседних клеток, образуя щелевые соединения. Недавние исследования показали существование связи между адгезионными и щелевыми соединениями, предполагая более высокий уровень координации, чем считалось ранее.
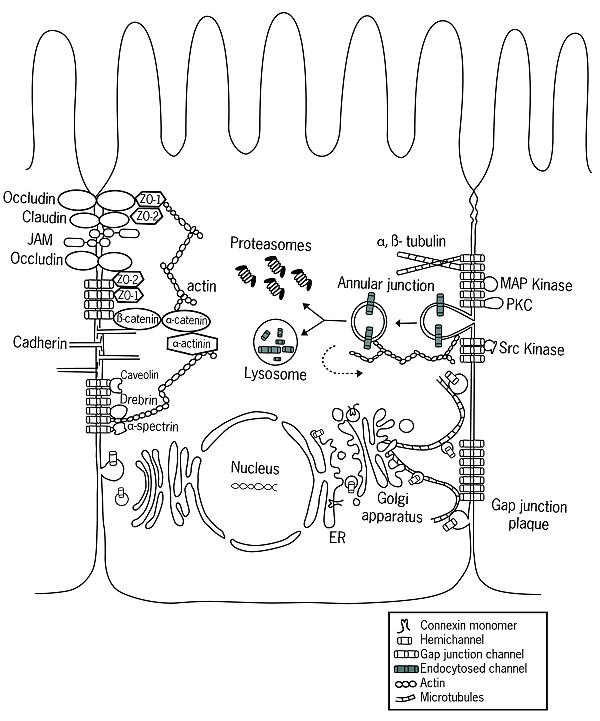 Жизненный цикл и белковые ассоциации коннексинов. Коннексины синтезируются на связанных с ЭР рибосомах и вставляются в ЭР котрансляционно. За этим следует олигомеризация между ER и сетью транс-Гольджи (в зависимости от типа коннексина) в коннексоны, которые затем доставляются к мембране через сеть актина или микротрубочек. Коннексоны также могут доставляться к плазматической мембране прямым переносом из грубого ER. После встраивания в мембрану коннексоны могут оставаться в виде полуканалов или стыковаться с совместимыми коннексонами на соседних клетках с образованием щелевых соединений. Вновь доставленные коннексоны добавляются к периферии предварительно сформированных щелевых соединений, в то время как центральный «старый» фрагмент щелевых соединений разрушается за счет интернализации двухмембранной структуры, называемой кольцевым соединением, в одну из двух клеток, где последующие лизосомные или протеасомные происходит деградация, или в некоторых случаях коннексоны возвращаются в мембрану (обозначено пунктирной стрелкой). В течение своего жизненного цикла коннексины связываются с различными белками, включая (1) компоненты цитоскелета, такие как микротрубочки, актин и актин-связывающие белки α- спектрин и дребрин, (2) соединительные молекулы включая компоненты слипчивого соединения, такие как кадгерины, α-катенин и β-катенин, а также компоненты плотного соединения, такие как ZO-1 и ZO-2, (3) ферменты, такие как киназы и фосфатазы, которые регулируют сборку, функцию и деградацию, и (4) другие белки, такие как кавеолин. Это изображение было подготовлено Hanaa Hariri для Dbouk et al., 2009.
Жизненный цикл и белковые ассоциации коннексинов. Коннексины синтезируются на связанных с ЭР рибосомах и вставляются в ЭР котрансляционно. За этим следует олигомеризация между ER и сетью транс-Гольджи (в зависимости от типа коннексина) в коннексоны, которые затем доставляются к мембране через сеть актина или микротрубочек. Коннексоны также могут доставляться к плазматической мембране прямым переносом из грубого ER. После встраивания в мембрану коннексоны могут оставаться в виде полуканалов или стыковаться с совместимыми коннексонами на соседних клетках с образованием щелевых соединений. Вновь доставленные коннексоны добавляются к периферии предварительно сформированных щелевых соединений, в то время как центральный «старый» фрагмент щелевых соединений разрушается за счет интернализации двухмембранной структуры, называемой кольцевым соединением, в одну из двух клеток, где последующие лизосомные или протеасомные происходит деградация, или в некоторых случаях коннексоны возвращаются в мембрану (обозначено пунктирной стрелкой). В течение своего жизненного цикла коннексины связываются с различными белками, включая (1) компоненты цитоскелета, такие как микротрубочки, актин и актин-связывающие белки α- спектрин и дребрин, (2) соединительные молекулы включая компоненты слипчивого соединения, такие как кадгерины, α-катенин и β-катенин, а также компоненты плотного соединения, такие как ZO-1 и ZO-2, (3) ферменты, такие как киназы и фосфатазы, которые регулируют сборку, функцию и деградацию, и (4) другие белки, такие как кавеолин. Это изображение было подготовлено Hanaa Hariri для Dbouk et al., 2009. Щелевые соединения коннексина обнаружены только у позвоночных, в то время как функционально аналогичная (но генетически неродственная) группа белков, иннексины, ответственны за щелевые соединения у видов беспозвоночных. Ортологи иннексина также были идентифицированы в хордовых, но они больше не способны образовывать щелевые соединения. Вместо этого каналы, образованные этими белками (называемые паннексинами ), действуют как очень большие трансмембранные поры, которые соединяют внутри- и внеклеточные компартменты.
Внутри ЦНС щелевые соединения обеспечивают электрическую связь между клетками-предшественниками, нейронами и глиальными клетками. Исследования показали, что при использовании мышей с нокаутом по коннексину , связывание клеток важно для передачи визуальных сигналов. В сетчатке уровни окружающего света влияют на сцепление клеток, обеспечиваемое каналами щелевого соединения, адаптируя зрительную функцию к различным условиям освещения. Сцепление клеток регулируется несколькими механизмами, включая экспрессию коннексина.
Decrock et al.. обсуждали многоуровневую платформу, с помощью которой коннексины и паннексины могут влиять на следующие клеточные функции в ткани: (1) каналы щелевых соединений коннексина (GJCs) обеспечивают прямую межклеточную коммуникацию малых молекул, (2) гемиканалы коннексина и каналы паннексина могут вносить вклад в аутокринные / паракринные пути передачи сигналов и (3) различные структурные домены эти белки обеспечивают независимые от каналов функции, такие как адгезия между клетками, взаимодействия с цитоскелетом и активация внутриклеточных сигнальных путей. Таким образом, коннексины и паннексины вносят многогранный вклад в развитие мозга и специфические процессы в нейроглио-сосудистой единице, включая синаптическую передачу и пластичность, глиальную передачу сигналов, вазомоторный контроль и целостность гематоэнцефалического барьера в зрелой ЦНС.
Различные коннексины могут проявлять различную специфичность в отношении растворенных веществ. Например, аденозин примерно в 12 раз лучше проходил через каналы, образованные Cx32, в то время как AMP и ADP проходил примерно в 8 раз лучше, а ATP более чем в 300 раз лучше, через каналы, образованные Сх43. Таким образом, добавление фосфата к аденозину, по-видимому, смещает его относительную проницаемость с каналов, образованных Cx32, на каналы, образованные Cx43. Это может иметь функциональные последствия, поскольку энергетический статус клетки может контролироваться посредством экспрессии коннексина и образования каналов.
Транспортная реакция, катализируемая щелевыми соединениями коннексина, имеет следующий вид:
| Коннексин | Ген | Расположение и функция |
|---|---|---|
| Cx43 | GJA1 | Экспрессируется в поверхность сосудистой сети с атеросклеротической бляшкой и активируется во время атеросклероза у мышей. Может иметь патологические эффекты. Также экспрессируется между клетками гранулезы, что необходимо для пролиферации. Обычно экспрессируется в астроцитах, также обнаруживается в большинстве астроцитом человека и в астроглиальном компоненте глионейрональных опухолей. Это также основной кардиальный коннексин, обнаруживаемый в основном в миокарде желудочков. Связано с окулодентодигитальной дисплазией. |
| Cx46 | GJA3 | |
| Cx37 | GJA4 | Индуцируется в гладких мышцах сосудов во время коронарного артериогенеза. Мутации Сх37 не смертельны. Образует щелевые контакты между ооцитами и клетками гранулезы и необходимы для выживания ооцитов. |
| Cx40 | GJA5 | избирательно экспрессируется в миоцитах предсердий. Отвечает за скоординированную электрическую активацию предсердий. |
| Cx33 | . (GJA6P) | Псевдоген у людей |
| Cx50 | GJA8 | Щелевые соединения между горизонтальными клетками A-типа у мышей и Retina кролика |
| Cx59 | GJA10 | |
| Cx62 | GJA10 | Human Cx62 соответствует Cx57 (мышь). Расположение в несущей аксон горизонтальной клетке B-типа в сетчатке кролика |
| Cx32 | GJB1 | Главный компонент периферического миелина. Мутации в человеческом гене вызывают Х-сцепленную болезнь Шарко-Мари-Тута, наследственную невропатию. В нормальном мозге человека CX32 экспрессируется в нейронах и олигодендроцитах. |
| Cx26 | GJB2 | Мутировал при синдроме Фохвинкеля, а также синдроме кератита-иктиоза-глухоты (KID). |
| Cx31 | GJB3 | Может быть связан с Erythrokeratodermia variabilis. |
| Cx30.3 | GJB4 | Fonseca et al. подтвердили экспрессию Cx30.3 в тимоцитах. Может быть связан с Erythrokeratodermia variabilis. |
| Cx31.1 | GJB5 | |
| Cx30 | GJB6 | Мутировал в синдром Клустона (гидротическая эктодермальная дисплазия) |
| Cx25 | GJB7 | |
| Cx45 | GJC1 / GJA7 | Эпителиальные клетки протоков поджелудочной железы человека. Атриовентрикулярный узел. |
| Cx47 | GJC2 / GJA12 | Экспрессируется в щелевых соединениях олигодентроцитов |
| Cx31.3 | GJC3 | Человеческий ортолог мышиного Cx29. Не известно об образовании щелевых контактов. |
| Cx36 | GJD2 / GJA9 | Функция бета-клеток поджелудочной железы, опосредующая высвобождение инсулина. Нейроны во всей центральной нервной системе, где они синхронизируют нервную активность. |
| Cx31.9 | GJD3 / GJC1 | |
| Cx39 | GJD4 | |
| Cx40.1 | GJD4 | |
| Cx23 | GJE1 |
Щелевые соединения необходимы для многих физиологических процессов, таких как скоординированная деполяризация сердечной мышцы, правильное эмбриональное развитие и управляемая реакция в микроциркуляторном русле. По этой причине делеция или мутация различных изоформ коннексина вызывает отличительные фенотипы и патологии. В то время как мутации в Сх43 в основном связаны с окулодентодигитальной дисплазией, мутации Сх47 связаны с Пелизеем-Мерцбахером -подобным заболеванием и лимфедемой. Мутации Сх40 в основном связаны с фибрилляцией предсердий. Мутации в Cx37 еще не описаны, но полиморфизмы в гене Cx37 вовлечены в развитие артериальных заболеваний.
На данный момент в этой статье используется контент из "1.A.24 Семейство коннексинов (коннексинов), образующих промежутки между соединениями ", которое лицензировано способом, допускающим повторное использование в соответствии с непортированной лицензией Creative Commons Attribution-ShareAlike 3.0, но не в соответствии с GFDL. Все соответствующие условия должны быть соблюдены.