
Войти
 Экстракт хлорофилла в спирте показан в белом свете (вверху) и в УФ-свете, вызывающем флуоресценцию (внизу).
Экстракт хлорофилла в спирте показан в белом свете (вверху) и в УФ-свете, вызывающем флуоресценцию (внизу). 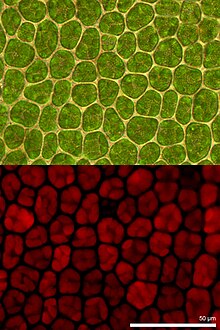 Микроскопические изображения лист мха из. Светлопольная микроскопия вверху и флуоресцентная микроскопия внизу. Красная флуоресценция исходит от хлорофилла в хлоропластах.
Микроскопические изображения лист мха из. Светлопольная микроскопия вверху и флуоресцентная микроскопия внизу. Красная флуоресценция исходит от хлорофилла в хлоропластах. Флуоресценция хлорофилла - это свет, переизлучаемый молекулами хлорофилла при возвращении из возбужденного состояния в невозбужденное. Он используется как индикатор преобразования фотосинтетической энергии у высших растений, водорослей и бактерий. Возбужденный хлорофилл рассеивает поглощенную световую энергию, приводя в действие фотосинтез (фотохимическое преобразование энергии), в виде тепла в нефотохимическом тушении или за счет испускания в виде флуоресцентного излучения. Поскольку эти процессы являются взаимодополняющими, анализ флуоресценции хлорофилла является важным инструментом в исследованиях растений с широким спектром применений.
При освещении адаптированного к темноте листа наблюдается быстрое увеличение флуоресценции от Фотосистемы II (ФСII), за которым следует медленное снижение. Впервые наблюдаемый Каутским и др. В 1932 г., это явление получило название эффекта Каутского. Этот переменный рост флуоресценции хлорофилла происходит из-за фотосистемы II. Флуоресценция фотосистемы I не переменная, а постоянная.
Увеличение флуоресценции происходит из-за того, что реакционные центры ФСII находятся в «закрытом» или химически восстановленном состоянии. Реакционные центры «закрываются», когда не могут принимать дальнейшие электроны. Это происходит, когда акцепторы электронов после ФСII еще не передали свои электроны следующему электронному носителю, поэтому они не могут принять другой электрон. Закрытые реакционные центры снижают общую фотохимическую эффективность и, таким образом, повышают уровень флуоресценции. При переводе листа из темноты в свет увеличивается доля закрытых реакционных центров ФСII, поэтому уровни флуоресценции увеличиваются в течение 1-2 секунд. Впоследствии флуоресценция уменьшается в течение нескольких минут. Это связано с; 1. большее «фотохимическое тушение», при котором электроны транспортируются от ФСII за счет ферментов, участвующих в фиксации углерода; и 2. большее «нефотохимическое тушение», при котором больше энергии преобразуется в тепло.
Обычно начальное измерение - это минимальный уровень флуоресценции, 
Чтобы использовать измерения флуоресценции хлорофилла для анализа фотосинтеза, исследователи должны различать фотохимическое тушение и нефотохимическое тушение (рассеивание тепла). Это достигается путем прекращения фотохимии, что позволяет исследователям измерять флуоресценцию только при наличии нефотохимического тушения. Чтобы уменьшить фотохимическое тушение до незначительного уровня, на лист направляют короткую вспышку света высокой интенсивности. Это временно закрывает все реакционные центры ФСII, что предотвращает передачу энергии ФСII нижележащим электронным носителям. Короткая вспышка не повлияет на нефотохимическое гашение. Во время вспышки флуоресценция достигает уровня, достигаемого в отсутствие какого-либо фотохимического гашения, известного как максимальная флуоресценция 
Эффективность фотохимического гашения (которая является косвенным показателем эффективности PSII) можно оценить, сравнив 












Пока
Флуоресценция хлорофилла, по-видимому, является мерой фотосинтеза, но это чрезмерное упрощение. Флуоресценция может измерять эффективность фотохимии ФСII, которая может использоваться для оценки скорости линейного переноса электронов путем умножения на интенсивность света. Однако исследователи обычно имеют в виду фиксацию углерода, когда говорят о фотосинтезе. Транспорт электронов и фиксация CO 2 могут хорошо коррелировать, но могут не коррелировать в полевых условиях из-за таких процессов, как фотодыхание, метаболизм азота и реакция Мелера.
Мощный метод исследования состоит в одновременном измерении флуоресценции хлорофилла и газообмена для получения полной картины реакции растений на окружающую среду. Один из методов состоит в одновременном измерении фиксации CO 2 и фотохимии PSII при разной интенсивности света в нефотодыхательных условиях. График фиксации CO 2 и фотохимии PSII указывает потребность в электронах на одну фиксированную молекулу CO 2. Исходя из этой оценки, можно оценить степень фотодыхания. Это было использовано для изучения значения фотодыхания как фотозащитного механизма во время засухи.
Флуоресцентный анализ также можно применять для понимания эффектов низких и высоких температур.
 ). В тех же листьях, используемых для измерений газообмена, параметры флуоресценции хлорофилла а (начальное, ; максимальное, и переменная, ) были измерены с использованием флуорометра. Результаты показали, что, несмотря на то, что виды-первопроходцы и лесные виды, населяющие разные среды обитания, оба показали одинаковую уязвимость к полуденному фотоингибированию на листьях, подвергшихся воздействию солнца.
). В тех же листьях, используемых для измерений газообмена, параметры флуоресценции хлорофилла а (начальное, ; максимальное, и переменная, ) были измерены с использованием флуорометра. Результаты показали, что, несмотря на то, что виды-первопроходцы и лесные виды, населяющие разные среды обитания, оба показали одинаковую уязвимость к полуденному фотоингибированию на листьях, подвергшихся воздействию солнца.Флуоресценция хлорофилла может измерять большинство типов стресс растений. Флуоресценция хлорофилла может использоваться в качестве индикатора стресса растений, поскольку стрессы окружающей среды, например экстремальные температуры, доступность света и воды могут снизить способность растения к нормальному метаболизму. Это может означать дисбаланс между поглощением световой энергии хлорофиллом и использованием энергии в фотосинтезе.
при полном солнечном свете у поздних сукцессионных видов, чем у пионерский вид. В целом, их результаты показывают, что виды-первопроходцы лучше проявляют себя при ярком солнечном свете, чем виды поздних сукцессий, что позволяет предположить, что растения-первопроходцы обладают большей потенциальной устойчивостью к фотоокислительному повреждению., и . На флуоресценцию хлорофилла листьев концентрация NaCl существенно не влияла, когда концентрация B была низкой. Когда B был увеличен, флуоресценция хлорофилла листьев уменьшалась в условиях солевого раствора. Можно сделать вывод, что комбинированное действие B и NaCl на малину вызывает токсический эффект по фотохимическим параметрам. Пример портативного многопараметрического флуориметра, который использует соотношение между хлорофиллом и флавонолами для определения дефицита азота в растениях
Пример портативного многопараметрического флуориметра, который использует соотношение между хлорофиллом и флавонолами для определения дефицита азота в растениях Из-за связь между содержанием хлорофилла и содержанием азота в листьях, флуорометры хлорофилла могут использоваться для обнаружения дефицита азота в растениях несколькими методами.
Основываясь на нескольких годах исследований и экспериментов, полифенолы может быть индикатором азотного статуса растения. Например, когда растение находится в оптимальных условиях, оно способствует своему первичному метаболизму и синтезирует белки (молекулы азота), содержащие хлорофилл, и небольшое количество флавонолов (вторичные соединения на основе углерода). С другой стороны, в случае нехватки азота мы будем наблюдать повышенное производство флавонолов растением.
NBI (индекс баланса азота) по Force-A позволяет оценить азотные условия в помещении. культивирование путем расчета отношения между хлорофиллом и флавонолами (в отношении выделения азота / углерода).
Гительсон (1999) утверждает: «Было обнаружено, что соотношение между флуоресценцией хлорофилла при 735 нм и диапазоном длин волн от 700 до 710 нм, F735 / F700 линейно пропорционально содержание хлорофилла (с коэффициентом детерминации r2, более 0,95), и, таким образом, это соотношение может использоваться в качестве точного индикатора содержания хлорофилла в листьях растений ».
 Изображение флуоресценции (значение Ft) адаксиальная поверхность листа
Изображение флуоресценции (значение Ft) адаксиальная поверхность листа Развитие флуорометров позволило флуоресцентному анализу хлорофилла стать обычным методом исследования растений. Флуоресцентный анализ хлорофилла произвел революцию в связи с изобретением метода амплитудно-импульсной модуляции (PAM) и появлением первого коммерческого флуориметра с модуляцией хлорофилла PAM-101 (Walz, Германия). Путем модуляции измерительного светового луча (импульсы микросекундного диапазона) и параллельного обнаружения возбужденной флуоресценции можно определить относительный выход флуоресценции (Ft) в присутствии окружающего света. Что особенно важно, это означает, что флуоресценцию хлорофилла можно измерить в полевых условиях даже при ярком солнечном свете.
Сегодня флуорометры хлорофилла предназначены для измерения множества различных механизмов растений. Протоколы измерений: F V/FMи OJIP измеряют эффективность образцов Photosystem II в обычном и известном состоянии адаптации к темноте. Эти протоколы полезны для измерения многих типов стресса растений. Протокол измерения ΔF / F M ’, или Y (II) Бернара Дженти, адаптированный к свету, является эффективным и чувствительным способом измерения образцов растений в условиях окружающего или искусственного освещения. Однако, поскольку значения Y (II) также изменяются в зависимости от интенсивности света, следует сравнивать образцы при одинаковой интенсивности света, если только световое напряжение не является фокусом измерения. Y (II) может быть более чувствительным к некоторым типам стресса растений, чем F V/FM, таким как тепловой стресс.
Также были разработаны другие протоколы измерения механизмов растений. Когда хлоропласт поглощает свет, часть световой энергии идет на фотохимию, часть идет на регулируемое рассеяние тепла, а часть идет на нерегулируемое рассеяние тепла. Существуют различные параметры измерения флуоресценции хлорофилла для измерения всех этих событий. В модели озера q L измеряет фотохимическое тушение, Y (NYO) измеряет регулируемое рассеяние тепла растением, а Y (NO) измеряет нерегулируемое рассеяние тепла. Более старый протокол гашения, называемый моделью лужи, использует q P для фотохимического гашения, q N для нефотохимического гашения как регулируемого, так и нерегулируемого рассеивания тепла и NPQ для оценки нефотохимического гашения. NPQ также был преобразован в модель озера математически.
Кроме того, были разработаны параметры q E и pNPQ для измерения цикла фотозащитного ксантофилла. q T - мера переходов состояний. q M представляет собой меру миграции хлоропластов, а q I представляет собой меру фотоингибирования растений.
При более низких уровнях актинического света NPQ = qE + qT + qI
При высоких уровнях актинической освещенности NPQ = qE + qM = qI
Некоторые флуорометры сконструированы так, чтобы их можно было носить в одной руке.
Последовательное дальнейшее развитие флюориметров для визуализации облегчает визуализацию пространственных неоднородностей фотосинтетической активности образцов. Эти неоднородности естественным образом возникают в листьях растений, например, во время роста, различных стрессов окружающей среды или инфекции патогенами. Таким образом, знание о неоднородности образцов важно для правильной интерпретации фотосинтетических характеристик образца растений. Высокопроизводительные флуориметрические системы визуализации предоставляют возможности для анализа отдельных клеток / отдельных хлоропластов, а также участков образцов, покрывающих целые листья или растения.
Методы, основанные на эффекте Каутского, не исчерпывают разнообразия методов обнаружения и оценки, основанных на флуоресценции хлорофилла. В частности, недавние достижения в области лазерно-индуцированной флуоресценции (LIF) также предоставляют возможность разработки достаточно компактных и эффективных датчиков для оценки фотофизиологического статуса и биомассы. Вместо измерения эволюции общего потока флуоресценции такие датчики регистрируют спектральную плотность этого потока, возбуждаемого мощными монохроматическими импульсами лазерного света с длительностью наносекунды. Не требуя 15-20-минутного периода адаптации к темноте (как в случае методов эффекта Каутского) и будучи способными возбуждать образец со значительного расстояния, датчики LIF могут обеспечить быструю и удаленную оценку.
Усовершенствованный флуориметр хлорофилла с непрерывным возбуждением