
Войти
 Хлорофилл отвечает за зеленый цвет многих растений и водоросли.
Хлорофилл отвечает за зеленый цвет многих растений и водоросли.  При наблюдении под микроскопом хлорофилл концентрируется внутри организмов в структурах, называемых хлоропластами, показанными здесь, сгруппированными внутри клеток растений.
При наблюдении под микроскопом хлорофилл концентрируется внутри организмов в структурах, называемых хлоропластами, показанными здесь, сгруппированными внутри клеток растений.  Растения воспринимаются как зеленые, потому что хлорофилл поглощает в основном синий и красный длина волны и отражает зеленый цвет.
Растения воспринимаются как зеленые, потому что хлорофилл поглощает в основном синий и красный длина волны и отражает зеленый цвет. 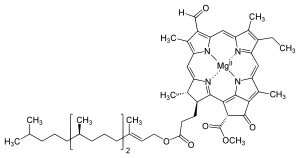 Существует несколько типов хлорофилла, но все они имеют общий хлорин магний лиганд, который составляет правую часть этой диаграммы.
Существует несколько типов хлорофилла, но все они имеют общий хлорин магний лиганд, который составляет правую часть этой диаграммы. Хлорофилл (также хлорофил ) представляет собой любой из нескольких связанных зеленых пигментов, обнаруженных в мезосомах цианобактерий и в хлоропласты водорослей и растений. Его название происходит от греческих слов χλωρός, khloros («бледно-зеленый») и φύλλον, phyllon («лист»). Хлорофилл необходим для фотосинтеза, позволяя растениям поглощать энергию света.
Хлорофиллы наиболее сильно поглощают свет в синей части электромагнитного спектра, а также в красной части. И наоборот, он плохо поглощает зеленые и почти зеленые части спектра, которые он отражает, создавая зеленый цвет хлорофилл-содержащих тканей. В фотосистемах зеленых растений существуют два типа хлорофилла: хлорофилл а и b.
Хлорофилл был впервые выделен и назван Жозефом Бьенэме Кавенту и Пьером Жозефом Пеллетье в 1817 году. магния в хлорофилле был обнаружен в 1906 году и явился первым обнаружением этого элемента в живых тканях.
После первоначальной работы, проделанной немецким химиком Ричардом Вильштеттером с 1905 по 1915 г., общая структура хлорофилла а была выяснена Гансом Фишером в 1940 г. К 1960 г., когда большая часть стереохимии хлорофилла Если было известно, Роберт Бернс Вудворд опубликовал полный синтез молекулы. В 1967 году последнее оставшееся стереохимическое разъяснение было завершено Яном Флемингом, а в 1990 году Вудворд и соавторы опубликовали обновленный синтез. Было объявлено, что хлорофилл f присутствует в цианобактерии и другие кислородсодержащие микроорганизмы, образующие строматолиты в 2010 г.; молекулярная формула C 55H70O6N4Mg и структура (2- формил ) -хлорофилла a были выведены на основе ЯМР, оптических и масс-спектров.
 Поглощение спектры свободного хлорофилла a (синий) и b (красный) в растворителе. Спектры молекул хлорофилла немного изменяются in vivo в зависимости от конкретных взаимодействий пигмент-белок.
Поглощение спектры свободного хлорофилла a (синий) и b (красный) в растворителе. Спектры молекул хлорофилла немного изменяются in vivo в зависимости от конкретных взаимодействий пигмент-белок. Хлорофилл жизненно важен для фотосинтеза, который позволяет растениям поглощать энергию света.
Молекулы хлорофилла расположены внутри и вокруг фотосистем, которые встроены в тилакоидные мембраны хлоропластов. В этих комплексах хлорофилл выполняет три функции. Функция подавляющего большинства хлорофилла (до нескольких сотен молекул на фотосистему) заключается в поглощении света. Сделав это, эти же центры выполняют свою вторую функцию: передачу этой световой энергии посредством резонансной передачи энергии определенной паре хлорофилла в реакционном центре фотосистем. Эта пара выполняет конечную функцию хлорофиллов, разделение зарядов, что приводит к биосинтезу. Двумя принятыми в настоящее время единицами фотосистемы являются фотосистема II и фотосистема I, которые имеют свои собственные отдельные реакционные центры, названные P680 и P700, соответственно. Эти центры названы в честь длины волны (в нанометров ) их максимума поглощения красного пика. Идентичность, функция и спектральные свойства типов хлорофилла в каждой фотосистеме различны и определяются друг другом и окружающей их белковой структурой. После экстракции из белка в растворитель (такой как ацетон или метанол ) эти пигменты хлорофилла могут быть разделены на хлорофилл a и хлорофилл b.
Функция реакционного центра хлорофилла - поглощать световую энергию и передавать ее другим частям фотосистемы. Поглощенная энергия фотона передается электрону в процессе, называемом разделением зарядов. Удаление электрона из хлорофилла - это реакция окисления. Хлорофилл отдает электрон высокой энергии ряду молекулярных промежуточных продуктов, называемых цепью переноса электронов. Затем заряженный реакционный центр хлорофилла (P680) восстанавливается до своего основного состояния, принимая электрон, оторванный от воды. Электрон, который восстанавливает P680, в конечном итоге возникает в результате окисления воды до O 2 и H через несколько промежуточных соединений. Эта реакция представляет собой то, как фотосинтезирующие организмы, такие как растения, производят газ O 2, и является источником практически всего O 2 в атмосфере Земли. Фотосистема I обычно работает последовательно с Фотосистемой II; таким образом, P700 Фотосистемы I обычно уменьшается, поскольку он принимает электрон через множество промежуточных продуктов в тилакоидной мембране, электронами, поступающими, в конечном счете, из Фотосистемы II. Однако реакции переноса электронов в тилакоидных мембранах являются сложными, и источники электронов, используемые для восстановления P700, могут варьироваться.
Электронный поток, производимый пигментами хлорофилла реакционного центра, используется для прокачки ионов H через тилакоидную мембрану, создавая хемиосмотический потенциал, используемый в основном для производства АТФ (запасенная химическая энергия) или для восстановления НАДФ до НАДФ. НАДФН - это универсальный агент, используемый для восстановления CO 2 до сахаров, а также для других биосинтетических реакций.
Хлорофилл-белковые комплексы с реакционным центром способны напрямую поглощать свет и выполнять процессы разделения зарядов без помощи других пигментов хлорофилла, но вероятность того, что это произойдет при данной интенсивности света, мала. Таким образом, все другие хлорофиллы в фотосистеме и белки антенного пигмента совместно поглощают световую энергию и направляют ее в реакционный центр. Помимо хлорофилла а, существуют другие пигменты, называемые дополнительными пигментами, которые встречаются в этих антенных комплексах пигмент-белок.
 Модель, заполняющая пространство молекулы хлорофилла а
Модель, заполняющая пространство молекулы хлорофилла а Типы хлорофиллов многочисленны, но все они определяются наличием пятого кольца за четырьмя пирролоподобными кольцами. Большинство хлорофиллов классифицируются как хлорины, которые являются восстановленными родственниками порфиринов (обнаружены в гемоглобине ). У них общий путь биосинтеза с порфиринами, включая предшественник уропорфириноген III. В отличие от гемов, которые содержат железо в центре тетрапиррольного кольца, хлорофиллы связывают магний. Для структур, изображенных в этой статье, некоторые из лигандов , прикрепленных к центру Mg, опущены для ясности. Хлориновое кольцо может иметь различные боковые цепи, обычно включая длинную цепь фитола. Наиболее распространенной формой у наземных растений является хлорофилл а.
Структуры хлорофиллов кратко описаны ниже:
| Хлорофилл a | Хлорофилл b | Хлорофилл c1 | Хлорофилл c2 | Хлорофилл d | Хлорофилл f | |
|---|---|---|---|---|---|---|
| Молекулярная формула | C55H72O5N4Mg | C55H70O6N4Mg | C35H30O5N4Mg | C35H28O5N4Mg | C54H70O6N4Mg | C55H70O6N4Mg |
| Группа C2 | -CH 3 | -CH 3 | -CH 3 | -CH 3 | -CH 3 | -CHO |
| группа C3 | -CH = CH 2 | -CH = CH 2 | -CH = CH 2 | -CH = CH 2 | -CHO | - CH = CH 2 |
| группа C7 | -CH 3 | -CHO | -CH 3 | -CH 3 | -CH 3 | -CH 3 |
| группа C8 | -CH 2CH3 | -CH 2CH3 | -CH 2CH3 | -CH = CH 2 | -CH 2CH3 | -CH 2CH3 |
| группа C17 | -CH 2CH2COO-Phytyl | -CH 2CH2COO-Phytyl | -CH = CHCOOH | -CH = CHCOOH | -CH 2CH2COO-Phytyl | -CH 2CH2COO-Phytyl |
| C17-C18 связь | Одинарная. (хлорин) | Одинарная. (хлорин) | Двойная. ( порфирин) | Двойной. (порфирин) | Одинарный. (хлорин) | Одинарный. (хлорин) |
| Встречаемость | Универсальный | В основном растения | Различные водоросли | Различные водоросли | Цианобакт eria | Цианобактерии |
хлорофилл a
хлорофилл b

хлорофилл c1

хлорофилл c2
хлорофилл d
хлорофилл f
 Хлорофилл образует темно-зеленые растворы в органических растворителях.
Хлорофилл образует темно-зеленые растворы в органических растворителях. Измерение поглощения света осложняется растворителем, используемым для извлечения хлорофилла из растительного материала, что влияет на полученные значения,
Измеряя поглощение света в красной и дальней красной областях, можно оценить концентрацию хлорофилла в листе.
Коэффициент флуоресцентного излучения можно использовать для измерения содержания хлорофилла. За счет возбуждения флуоресценции хлорофилла а на более низкой длине волны соотношение испускания флуоресценции хлорофилла при 705 ± 10 нм и 735 ± 10 нм может обеспечить линейную зависимость содержания хлорофилла по сравнению с химическим тестированием. Отношение F 735 / F 700 обеспечивало значение корреляции r 0,96 по сравнению с химическим тестированием в диапазоне от 41 мг · м до 675 мг · м. Гительсон также разработал формулу для прямого считывания содержания хлорофилла в мг / м3. Формула обеспечивает надежный метод измерения содержания хлорофилла от 41 мг м до 675 мг м со значением корреляции 0 0,95.
У некоторых растений хлорофилл получают из глутамат и синтезируется по разветвленному биосинтетическому пути, который является общим с гемом и сирогемом. Хлорофиллсинтаза является ферментом который завершает биосинтез хлорофилла a, катализируя реакцию EC 2.5.1.62
 хлорофилла a + дифосфат
хлорофилла a + дифосфатЭто образует сложный эфир группы карбоновой кислоты в хлорофиллиде а с 20-углеродным дитерпеновым спиртом фитолом. Хлорофилл b вырабатывается тем же ферментом, который действует на хлорофиллид b.
У растений покрытосеменных более поздние стадии биосинтетического пути зависят от света, и такие растения бледные (этиолированные ) при выращивании в темноте. Несосудистые растения и зеленые водоросли имеют дополнительный светонезависимый фермент и становятся зелеными даже в темноте.
Хлорофилл сам по себе является связаны с белками и могут передавать поглощенную энергию в нужном направлении. Протохлорофиллид, один из биосинтетических промежуточных продуктов, встречается в основном в свободной форме и в условиях освещения действует как фотосенсибилизатор, образуя высокотоксичные свободные радикалы. Следовательно, растениям необходим эффективный механизм регулирования количества этого предшественника хлорофилла. У покрытосеменных это делается на стадии аминолевулиновой кислоты (ALA), одного из промежуточных соединений в пути биосинтеза. В растениях, которые питаются АЛК, накапливаются высокие и токсичные уровни протохлорофиллида; то же самое происходит и с мутантами с поврежденной регуляторной системой.
Процесс старения растений включает деградацию хлорофилла: например, фермент хлорофиллаза (EC 3.1.1.14 ) гидролизует боковую цепь фитила, чтобы обратить реакцию, в которой хлорофиллы биосинтезируются из хлорофиллида a или b. Поскольку хлорофиллид a может быть преобразован в хлорофиллид b, а последний может быть повторно этерифицирован до хлорофилла b, эти процессы позволяют циклически переключаться между хлорофиллами a и b. Более того, хлорофилл b может быть напрямую восстановлен (через 7-гидроксихлорофилл а) обратно до хлорофилла а, завершая цикл. На более поздних стадиях старения хлорофиллиды превращаются в группу бесцветных тетрапирролов, известных как нефлуоресцентные катаболиты хлорофилла (NCC), с общей структурой:
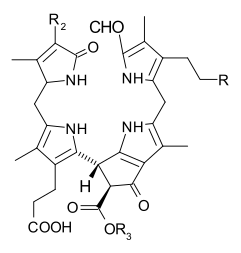
Эти соединения также были обнаружены в созревающих плодах, и они дают характерные особенности. осенняя окраска - лиственные растения.
Хлороз - это состояние, при котором листья производят недостаточно хлорофилл, делая их желтыми. Хлороз может быть вызван дефицитом питательных веществ железа, называемым хлорозом железа, или недостатком магния или азота. PH почвы иногда играет роль в хлорозе, вызванном питательными веществами; многие растения приспособлены к росту в почвах с определенным уровнем pH, и от этого может зависеть их способность поглощать питательные вещества из почвы. Хлороз также может быть вызван патогенами, включая вирусы, бактерии и грибковые инфекции, или насекомыми, сосущими сок.
 Наложение спектров хлорофилла a и b с оенином (мальвидин 30 глюкозид), типичный антоцианидин, показывая, что, хотя хлорофиллы поглощают в синей и желтой / красной частях видимого спектра, оенин поглощается в основном в зеленой части спектра, где хлорофиллы не поглощаются. совсем не впитываются.
Наложение спектров хлорофилла a и b с оенином (мальвидин 30 глюкозид), типичный антоцианидин, показывая, что, хотя хлорофиллы поглощают в синей и желтой / красной частях видимого спектра, оенин поглощается в основном в зеленой части спектра, где хлорофиллы не поглощаются. совсем не впитываются. Антоцианы - это другие растительные пигменты. Характер поглощения, отвечающий за красный цвет антоцианов, может быть дополнительным к таковому у зеленого хлорофилла в фотосинтетически активных тканях, таких как молодые листья Quercus coccifera. Он может защитить листья от нападений пожирателей растений, которых может привлечь зеленый цвет.
Карты хлорофилла показывают миллиграммы хлорофилла на кубический метр морской воды каждый месяц. Места, где количество хлорофилла было очень низким, что указывает на очень низкую численность фитопланктона, выделены синим цветом. Места, где концентрация хлорофилла была высокой, что означает рост многих фитопланктона, окрашены в желтый цвет. Наблюдения производятся спектрорадиометром среднего разрешения (MODIS) на спутнике НАСА Aqua. Земля темно-серая, а места, где MODIS не мог собрать данные из-за морского льда, полярной темноты или облаков, светло-серые. Самые высокие концентрации хлорофилла там, где крошечные обитающие на поверхности океанские растения процветают, находятся в холодных полярных водах или в местах, где океанские течения выносят холодную воду на поверхность, например, вокруг экватора и вдоль берегов континентов.. Фитопланктон стимулируется не самой холодной водой. Вместо этого низкие температуры часто являются признаком того, что вода поднялась на поверхность из глубин океана, неся питательные вещества, накопившиеся с течением времени. В полярных водах питательные вещества накапливаются в поверхностных водах в темные зимние месяцы, когда растения не могут расти. Когда весной и летом возвращается солнечный свет, растения цветут в высоких концентрациях.
Синтетический хлорофилл зарегистрирован как краситель для пищевых добавок, и его номер E это E140. Повара используют хлорофилл для окрашивания в зеленый цвет различных продуктов и напитков, например пасты и спиртных напитков. Абсент приобретает зеленый цвет естественным образом за счет хлорофилла, поступающего через большое количество различных трав, используемых при его производстве. Хлорофилл не растворяется в воде, и его сначала смешивают с небольшим количеством растительного масла для получения желаемого раствора.
Исследование 2002 года показало, что " листья, подвергнутые воздействию сильного света, содержали деградированные основные антенные белки, в отличие от тех, которые хранятся в темноте, что согласуется с исследованиями освещения изолированных белков ». Это показалось авторам подтверждением гипотезы о том, что «активные формы кислорода играют роль in vivo » в краткосрочном поведении растений.
| На Викискладе есть материалы, связанные с хлорофиллом. |