Войти
Кластер дифференцировки 40, CD40 - костимулирующий белок, обнаруженный на антигенпрезентирующих клетках и необходимый для их активация. Связывание CD154 (CD40L ) на THклетках с CD40 активирует антигенпрезентирующие клетки и вызывает множество последующих эффектов.
Дефицит может вызвать синдром гипер-IgM типа 3.
Белковый рецептор, кодируемый этим геном, является членом суперсемейства TNF-рецепторов. Было обнаружено, что этот рецептор играет важную роль в опосредовании широкого спектра иммунных и воспалительных реакций, включая Т-клеточно-зависимое переключение класса иммуноглобулинов, развитие В-клеток памяти и формирование зародышевого центра. AT-hook фактор транскрипции AKNA, как сообщается, скоординированно регулирует экспрессию этого рецептора и его лиганда, что может быть важно для гомотипических клеточных взаимодействий. Адаптерные белки фактора, связанные с TNFR-рецептором TRAF1, TRAF2, TRAF6 и, возможно, TRAF5 взаимодействуют с этим рецептором, и служат медиаторами сигнала. трансдукция. Обнаружено, что взаимодействие этого рецептора и его лиганда необходимо для активации микроглии, вызванной бета-амилоидом, и, таким образом, считается ранним событием в патогенезе болезни Альцгеймера. Сообщалось о двух альтернативно сплайсированных вариантах транскрипта этого гена, кодирующих разные изоформы.
В макрофаге первичным сигналом для активации является IFN-γ из Th1 типа CD4 Т-клеток. Вторичным сигналом является CD40L (CD154) на Т-клетке, которая связывает CD40 на поверхности клетки макрофага. В результате макрофаг экспрессирует больше рецепторов CD40 и TNF на своей поверхности, что помогает повысить уровень активации. Увеличение активации приводит к индукции сильнодействующих микробицидных веществ в макрофагах, включая активные формы кислорода и оксид азота, что приводит к уничтожению проглоченного микроба.
B-клетка может представлять антигены. Если активированная Т-клетка распознает пептид, представленный В-клеткой, CD40L на Т-клетке связывается с рецептором CD40 В-клетки, вызывая активацию В-клеток. Т-клетки также продуцируют ИЛ-2, который напрямую влияет на В-клетки. В результате этой чистой стимуляции В-клетка может подвергаться делению, переключению изотипа антитела и дифференцировке в плазматические клетки. Конечным результатом является В-клетка, которая способна массово продуцировать специфические антитела против антигенной мишени. Ранние доказательства этих эффектов заключались в том, что у мышей с дефицитом CD40 или CD154 наблюдается незначительное переключение классов или формирование зародышевого центра, а иммунные ответы сильно подавляются.
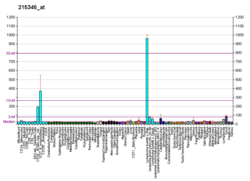
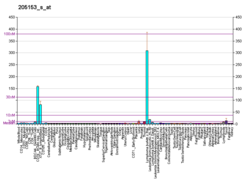
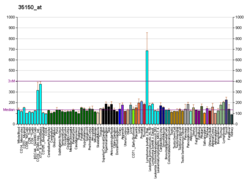
Экспрессия CD40 разнообразна. CD40 конститутивно экспрессируется антигенпрезентирующими клетками, включая дендритные клетки, В-клетки и макрофаги. Он также может экспрессироваться эндотелиальными клетками, клетками гладких мышц, фибробластами и эпителиальными клетками. В соответствии с его широко распространенной экспрессией на нормальных клетках, CD40 также экспрессируется на широком спектре опухолевых клеток, включая неходжкинские лимфомы и лимфомы Ходжкина, миелому и некоторые карциномы, включая носоглотку, мочевой пузырь, шейка матки, почка и яичник. CD40 также экспрессируется на предшественниках B-клеток в костном мозге, и есть некоторые свидетельства того, что взаимодействия CD40-CD154 могут играть роль в контроле B-клеточного гематопоэза.
CD40 (белок) было показано, что взаимодействует с TRAF2, TRAF3, TRAF6, TRAF5 и TTRAP.
Молекула CD40 является потенциальной мишенью для иммунотерапии рака. Существует ряд завершенных и продолжающихся клинических испытаний, в которых агонистические анти-CD40 моноклональные антитела используются для активации противоопухолевого Т-клеточного ответа посредством активации дендритных клеток.





 ..
..  ..
..