
Войти
Эволюция - это изменение характеристик биологических популяций в последовательных поколениях. Эти характеристики представляют собой выражения и генов, которые передаются от родителей к потомству во время воспроизводства. Различные характеристики имеют тенденцию влюбой данной популяции в результате мутации, генетической рекомбинации и других источников генетической изменчивости. Эволюция происходит, когда на эту вариацию воздействуют эволюционные процессы, такие как естественный отбор (включая половой отбор ) и генетический дрейф, в результате чего характеристики становятся более частыми или редкими в пределах населения. Именно этот процесс эволюции привел к биоразнообразию на всехуровнях биологической организации, включая уровни видов, отдельных организмов и молекулы.
научная теория эволюции путем естественного отбора независимо от системы Чарльзом Дарвином и Альфредом Расселом Уоллесом в середине XIX века. века и подробно изложен в книге Дарвина О происхождении видов. Эволюция путем естественного отбора впервые проведена наблюдением. Далее следуют три наблюдаемых факта о живыхорганизмах: (1) признаки различаются у разных людей в зависимости от их морфологии, физиологии и поведения (фенотипические вариации ), ( 2) разные черты дают разные показатели выживаемости и воспроизводимость (разная приспособленность ) и (3) черты могут передаваться из поколения в поколение (наследуемость приспособления). Таким образом, в рамках системы поколений члены пополняются с большей вероятностью замены потомками, которыепозволяют им выживать и воспроизводить в соответствующих средах. В начале 20 века другие конкурирующие идеи эволюции, такие как мутационизм и ортогенез, были опровергнуты как современный синтез согласовал дарвиновскую эволюцию с классической генетикой, которая установила, что адаптивная эволюция вызвана естественным отбором, воздействующим на менделевскую генетическую изменчивость.
Всяжизнь на Земле разделяет последнего всеобщего общего предка (LUCA), который жил примерно 3,5–3,8 миллиарда лет назад. палеонтологическая летопись включает в себя прогрессию от раннего биогенного графита к микробному мату окаменелостям и окаменелым многоклеточным организмом. Существующие модели биоразнообразия были сформированы другими видами образованиями новых видов (видообразование ), изменениями внутри видов (анагенез ) и утратой видов (исчезновение ) на протяжении всего периода эволюционная история жизни на Земле. Морфологические и биохимические признаки более схожи у видов, имеющих более общего предка, и могут быть для реконструкции филогенетических деревьев.
Эволюционные биологические Мы изучили различные аспекты эволюции, формируя и проверяя гипотезы, а также конструируя теории, основанные на критерии с продолжением поля или лаборатории и данных, полученные методы математической и теоретической биологии. Их открытия повлияли не только на развитие биологии, но и на множестве других научных и промышленных предприятий, включая сельское хозяйство, медицину и информатику.
 Лукреций
Лукреций  Альфред Рассел Уоллес
Альфред Рассел Уоллес  Томас Роберт Мальтус
Томас Роберт Мальтус  В 1842 году Чарльз Дарвин написал свой первый набросок Происхождение видов.
В 1842 году Чарльз Дарвин написал свой первый набросок Происхождение видов.Предположение, что один тип системы может происходить от другого типа восходит к другим из первых досократов греческих философов, таких как Анаксимандр и Эмпедокл. Такие предложения сохранились до римских времен. по и философ Лукреций последовали примеру Эмпедокла в его шедевре De rerum natura (О природе вещей).
В отличие от этих материалистических взглядов, аристотелианство рассматривало все естественные вещи как актуализации фиксированных природных возможностей, известных возможностей, возможности, возможности, как формы. Это было частью средневекового телеологического понимания природы, в котором все вещидолжны играть определенную роль в божественном космическом порядке. Вариации этой идеи стали стандартным пониманием Средневековье и были интегрированы в христианскую науку, но Аристотель не требовал, чтобы реальные типы организма всегда соответствовали один-к-одному точному метафизические и конкретно метафизические формы., могут новые живые существ.
В 17 веке появился новый метод из современная наука отвергла аристотелевский подход. Он искал объяснения природных явлений в терминах физических явлений, которые были одинаковы для всех видимых вещей и не требовали существования каких-либо фиксированных природных или космического порядка. Однако этот новый подход медленно укоренился в биологических науках, последнем использовании природных ресурсов. Джон Рэй применимо к одному из более общих терминов для фиксированных природных типов, «видов», к типам растений и животных, ноон строго определил каждый тип живых существ как вид и предположил, что каждый вид может быть определен чертами, которые сохраняются из поколения в поколение. Биологическая классификация, введенная Карлом Линнеем в 1735 году, явно признала иерархическую природу взаимоотношений видов, но по-прежнему рассматривала виды как фиксированные согласно божественному плану.
Другие натуралисты в это время размышляли об эволюционном изменении видов с течением времени всоответствии с законами природы. В 1751 году Пьер Луи Мопертюи писал о естественных модификациях, происходящих во время воспроизводства и накапливающихся в течение многих поколений, чтобы повторять новые виды. Жорж-Луи Леклерк, граф де Бюффон предположил, что виды могут вырождаться в разные виды. организмов, и Эразм Дарвин предположил, что все теплокровные животные происходили от одного микроорганизма (или «нити»). Первой полноценной эволюционной схемой была теория «трансмутации» Жана-Батиста Ламарка 1809 года, которая предусматривает спонтанное зарождение, непрерывно производящее простые простые формы жизни, которые развиваются все более сложными в параллельных линиях. с присущей им прогрессивной тенденцией. (Последний процесс позже был назван ламаркизмом.) Эти идеи были осуждены авторитетными натуралистами как предположение, не имеющее эмпирической поддержки. В частности, Жорж Кювье настаивал на том, что видыне связаны между собой и фиксированы, а их сходство отражает божественный замысел для функциональных нужд. Тем временем идеи доброжелательного замысла Рэя были развиты Уильямом Пейли в Естественная теология или свидетельства существования и атрибуты Божества (1802), предлагались сложные адаптации как божество божественного замысла. восхищался Чарльз Дарвин.
Решающий разрыв с концепцией постоянных типов различных типов биологиипроизошел с теориейциями посредством естественного отбора, был разработан Дарвином в терминах популяций. Дарвин использовал выражение «происхождение с модификацией », а не «эволюция». Частично находясь под областью «Очерка о принципах народонаселения (1798) Томаса Роберта Мальтуса, Дарвин заметил, что рост населения приведет к« борьбе за существование », в которых преобладают благоприятные вариации. другие погибли. В каждом поколении многие потомки недоживают до эпохи воспроизводства из-за ограниченных ресурсов. Это могло бы разнообразие растений и животных от общей родословной природы одинаково для всех типов организма. Дарвин разработал свою теорию «естественного отбора» с 1838 года и писал свою «большую книгу» по этому вопросу, когда Альфред Рассел Уоллес прислал ему версию практически той же теории в 1858 году. Их отдельные статьи были представлены вместе в 1858 году на собрании Лондонского общества Линнея.В конце 1859 года Дарвин опубликовал свой «реферат» под названием «Происхождение видов», который подробно объяснил естественный отбор таким образом, что привел к все более широкому признанию концепции эволюции Дарвина за счет альтернативные теории. Томас Генри Хаксли применил идеи Дарвина к людям, используя палеонтологию и сравнительную анатомию, чтобы предоставить убедительные доказательства того, что люди и обезьяны имели общуюродословную.. Некоторых это беспокоило, поскольку это означало, что люди не занимают особого места во вселенной .
Механизмы репродуктивной наследственности и происхождение новых черт оставались загадкой. С этой целью Дарвин разработал свою предварительную теорию пангенезиса. В 1865 г. Грегор Мендель сообщил, что черты наследуются таким образом через независимый набор и сегрегацию элементов (позже как известные гены).Законы наследования Менделя в итоге вытеснили часть теории пангенезиса Дарвина. Август Вейсман провел важное различие между зародышевыми клетками, дающими начало гаметам (такими как сперматозоиды и яйцеклетки ) и соматические клетки тела, демонстрируя, что наследственность передается только по зародышевой линии. Гуго де Фрис соединил теорию пангезиса Дарвина с различием клеток зародыша / сомы Вейсмана и предположил, чтопангены Дарвина концентрируются в ядре клетки и при экспрессии могут перемещаться в цитоплазму, чтобы изменить клетки . Де Фрис был также одним из исследователей, которые сделали работу Менделя широко известной, используемой, что менделевские черты соответствуют наследственной шкале вдоль зародышевой линии. Чтобы объяснить, как создать новые варианты, де Фрис разработал теорию мутаций, которая привела к временному разрыву между теми, кто принимаетдарвиновскую эволюцию, и биометристами, которые были союзниками де Вриза. В 1930-х годах пионеры в области популяционной генетики, такие как Рональд Фишер, Сьюолл Райт и Дж. Б. С. Холдейн заложил основы эволюции на прочную статистическую философию. Таким образом было разрешено ложное противоречие между теорией Дарвина, генетическими мутациями и менделевской наследственностью.
В 1920-х и 1930-х годах теназываемый современный синтез соединил естественный отбор и популяционную генетику, основанную на менделевской наследственности, в единую теорию, которая применима в целом к любой отрасли биологии. Современный синтез объяснил закономерности, наблюдаемые у разных видов в популяциях, через переходы в ископаемые в палеонтологии и сложные клеточные механизмы в биологии. Публикация структуры ДНК Джеймсом Уотсоном и Фрэнсисом Криком приучастии Розалинды Франклин в 1953 году использовала физический механизм наследования.. Молекулярная биология улучшила понимание взаимосвязи между генотипом и фенотипом. Были также достигнуты успехи в филогенетической систематике, отображающих переходных элементов в сравнительной и проверяемой системе использования и использовании систематике деревьев. В 1973 году биолог-эволюционист Феодосий Добжанский написал, что «ничто в биологии не имеет смысла, кроме как в свете эволюции », потому что это пролило свет на взаимосвязь между тем, что сначала предполагаемые несвязанные фактами в естественной в связную пояснительную совокупность знаний, которые предсказывает многие факты о жизни на планете.
С тех пор современный синтез был расширен для объяснения биологических явлений по всей и интегративной шкале биологической иерархии, от генов к видум. Однорасширение, известное как эволюционная биология развития и неофициально называемое «evo-DevO», выделяющееся как эволюция поколениями () влияет на паттерны изменений внутри отдельных организмов (развитие ). С начала 21 века и в свете открытий, сделанных в последние десятилетия, некоторые биологи выступали за расширенный эволюционный синтез, который мог бы объяснять эффекты негенетических способов наследования, таких как эпигенетика, дополняющиеэффекты, экологическая наследственность и культурная наследственность и эволюционируемость.
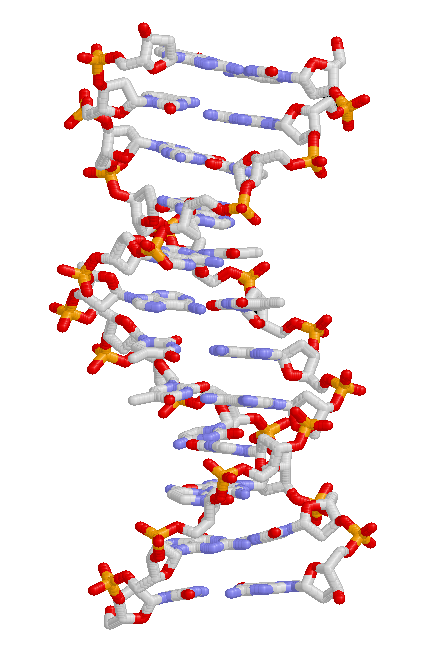 структура ДНК. Основания находятся в центре окруженных фосфатно-сахарными цепями двойная спирали.
структура ДНК. Основания находятся в центре окруженных фосфатно-сахарными цепями двойная спирали.Эволюция организма происходит через изменения наследственных признаков - унаследованных характеристик организма. Например, у людей цвет глаз является унаследованной характеристикой, и человек может унаследовать «черту карихглаз» от одного из своих родителей. Унаследованные признаки контролируются генами, и полный набор генов в геноме (генетический материал) организма называется его генотипом.
Полный набор наблюдаемых признаков, влияющих на структуру и поведение организма, называется его фенотипом. Эти черты возникают из-за взаимодействия его генотипа с окружающей средой. В результате многие аспекты фенотипа организма не передаются по наследству. Например, загорелая кожа возникает врезультате взаимодействия генотипа человека и солнечного света; таким образом, загар не передается детям людей. Однако некоторые люди загорают легче, чем другие из различных генотипов; Ярким примером являются люди с унаследованной чертой альбинизма, совсем не загорающая и очень чувствительная к солнечным ожогам.
Наследственные черты передаются от поколения к поколению через ДНК, молекула, кодирующая генетическая информацию. ДНК представляет собой длинный биополимер, состоящий из четырех типов оснований. Последовательность оснований в молекуле ДНК обеспечивает генетическую аналогично систему, производящую предложение. Перед делением клетки ДНК копируется, каждая из двух полученных клеток унаследует последовательность ДНК. Части молекулы ДНК, которые определяют одну функциональную единицу, называются генами; разные гены имеют разные наборы основ. Внутри клетки длинные цепи ДНК образуют конденсированные структуры, называемые хромосомами. Конкретное расположение ДНК в хромосоме известно как локус. Если последовательность ДНК в локусе у разных людей различается, разные формы этой системы называются аллелями. Последовательности ДНК могут изменяться в результате мутаций, производя новые аллели. Если в гене происходит мутация, новый аллель может повлиять на признак, который контролирует ген, изменяя фенотип организма. Однако, хотя это простое соответствие между аллелем и признаком работает в некоторых случаях,основные признаки являются более сложными и контролируются локусами количественных признаков (множественные взаимодействующие гены).
Недавние открытия подтвердили важные примеры наследственных изменений, объяснить изменениями последовательности нуклеотидов в ДНК. Эти явления классифицируются как эпигенетические системы наследования. метилирование ДНК маркировка хроматина, самоподдерживающиеся метаболические петли, сайленсинг геновпосредством интерференции РНК и трехмерная конформация белков (таких как прионы ) - это области, которых были обнаружены системы эпигенетического наследования на уровне организма. Биологи развития предполагают, что сложные взаимодействия в генетических животных и коммуникации между клетками приводят к наследственным вариациям, которые могут лежать в основе некоторых механизмов пластичности развития и канализации.Наследственность также может проявляться в еще более крупных масштабах. Например, экологическая наследственность в процессе строительства ниши определяет регулярную и регулярную деятельность систем в окружающей среде. Это порождает унаследованные эффекты, которые изменяют и возвращают режим выбора поколений. Потомки наследуют гены и характеристики окружающей среды, порожденные экологическими действиями предков. Другие примеры наследуемости в эволюции, которые не находятся под прямымконтролем генов, включают наследование культурных признаков и симбиогенез.
 Белая моль бёдер
Белая моль бёдер  Черная морфа в эволюция берёзовой моли
Черная морфа в эволюция берёзовой моли Фенотип отдельного организма является результатом его генотипа, так и условий окружающей среды, в которой он жил. Существенная часть фенотипической изменчивости в популяции вызвана генотипической изменчивостью. Современный эволюционный генератор определяет эволюцию как изменение во времени этой генетическойвариации. Частота одного конкретного аллеля станет более или менее предпочтительной по сравнению с другими формами этого гена. Вариация исчезает, когда новый аллель достигает точки фиксации - когда он либо исчезает из популяции, либо полностью заменяет предковый аллель.
Естественный отбор вызовет эволюцию только в том случае, если их достаточно генетическая изменчивость в популяции. До открытия менделевской генетики одной из распространенных гипотез было смешанное наследование. При смешанном наследовании генетическая изменчивость будет быстро утрачена, что сделает эволюцию путем отбора естественного маловероятной. Принцип Харди-Вайнберга обеспечивает решение, как поддерживается вариация в популяции с менделевским наследованием. Частоты аллелей (вариаций в гене) останутся постоянными в отсутствии отбора, мутаций, миграции и генетического дрейфа.
Вариация возникновения из-за мутаций в геноме, перетасовки генов посредствомполового размножения и миграция между популяциями (поток генов ). Несмотря на постоянное введение новых вариаций через мутации и поток генов, большая часть генома вида идентична у всех особей этого вида. Однако даже небольшие различия в генотипах могут привести к резким различиям в фенотипах: например, шимпанзе и люди различаются лишь примерно в 5% геномов.
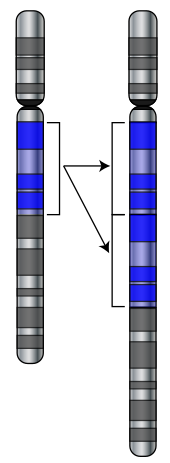 Дублирование части хромосома
Дублирование части хромосома Мутации - это изменения в последовательной ДНК геномаклетки. Когда проходят мутации, они могут улучшить продукт гена, или препятствовать функционированию гена, или не иметь эффекта. На основании на мухе Drosophila melanogaster было высказано предположение, что если мутация изменяет белок, продуцируемый геном, это, вероятно, будет вредным, так как около 70% этих мутаций будут иметь повреждающие эффекты, и остальная часть либо нейтральным, либо слабо полезна.
Мутации включения дублирование больших участковхромосомы (обычно в результате генетической рекомбинации ), что может привести к появлению дополнительных копий хромосомы. ген в геном. Дополнительные копии генов основного сырья, необходимого для развития новых генов. Это важно, потому что большинство новых генов эволюционируют в рамках семейств генов из уже существующих генов, имеющих общих предков. Например, человеческий глаз использует четыре гена для создания структур, воспринимающих свет: три для цветового зрения и один для ночного видения ; все четыре поколения от одного предкового гена.
Новые гены могут быть созданы из предкового гена, когда дублирующая копия мутирует и приобретает новую функцию. Этот процесс становится проще после дублирования гена, поскольку он увеличивает избыточность системы; один ген в паре может приобретать новую функцию, в то время как другая копия выполняет свою первоначальную функцию. Другие типы мутаций могут дажегенерировать совершенно новые гены из ранее некодирующей ДНК.
Генерация новых генов может также выполнить дупликацию небольших частей нескольких генов, при этом эти фрагменты рекомбинируют с образованием новых комбинаций с новыми функциями. Когда новые гены собираются из перетасов ранее существовавших частей, домены новых смешанных функций, которые можно использовать вместе для создания комбинаций с новыми и сложными функциями. Например, поликетидсинтазы - это большиеферменты, которые производят антибиотики ; они до ста независимых доменов, каждый из которых использует одну стадию общего процесса, например, стадию на конвейере.
В бесполых организмах, гены наследуются вместе или связаны, поскольку они не могут смешиваться с генами других организмов во время воспроизводства. Напротив, потомство половых содержит случайные смеси хромосом своих родителей, полученные путемнезависимогоотбора. В родственном процессе, называемом гомологичной рекомбинацией, половые организмы обмениваются ДНК между двумя совпадающими хромосомами. Рекомбинация и повторная сортировка не изменяют частоты аллелей, а вместо этого изменяют, какие аллели связаны друг с другом, производя потомство с новыми комбинациями аллелей. Секс обычно увеличивает генетическую изменчивость и может увеличить скорость эволюции.
 Эта диаграмма показывает двойную цену секса. Если каждая особьбудет даватьодно и то же количество потомков (два), (a) популяция полового останется того же размера в каждом поколении, где (b) популяция бесполого размножения удваивается в размере каждого поколения.
Эта диаграмма показывает двойную цену секса. Если каждая особьбудет даватьодно и то же количество потомков (два), (a) популяция полового останется того же размера в каждом поколении, где (b) популяция бесполого размножения удваивается в размере каждого поколения. Двукратная цена секса была впервые описана Джоном Мейнардом Смитом. Первая цена заключается в том, что у видов с половым диморфизмом только один из двух полов может рожать детенышей. (Эта стоимость не распространяется на виды-гермафродиты, такие какбольшинство различныхбеспозвоночных.) Вторая цена заключается в том, любой человек, который размножается половым путем, может передать только 50% своих генов любому отдельному потомству, даже с меньшим передается каждым новым поколением. Тем не менее, половое размножение наиболее распространенным средством размножения среди эукариот и многоклеточных организмов. Гипотеза Красной Королевы использовалась для объяснения значения полового размножения как средства,обеспечивающего непрерывнуюэволюцию и адаптацию в ответ на коэволюцию другими видами в постоянно меняющейся среде. 87>
Поток генов - это обмен генами между популяциями и между видами. Следовательно, это может быть новый для популяции вид источник изменчивости. Поток генов может быть вызван перемещением особей между устойчивыми популяциями или перемещением пыльцы между устойчивыми к тяжелым металлам и тяжелыми -металлочувствительными популяции травм.
Перенос геновмежду видами включает образование гибридных организмов и горизонтальный перенос генов. Горизонтальный перенос генов - это перенос генетического материала от одного организма к другому, не являющемуся его потомством; это наиболее распространено среди бактерий. В медицине это способствует распространению устойчивости к антибиотикам, поскольку одна бактерия приобретает гены устойчивости, она может быстро передать их другим видам.Произошел горизонтальныйперенос генов от бактерий к эукариотам, таким как дрожжи Saccharomyces cerevisiae и долгоносик бобов адзуки Callosobruchus chinensis. Примером крупномасштабного переноса являются эукариотические бделлоидные коловратки, которые получили ряд генов от бактерий, грибов и растений. Вирусы также могут нести ДНК между организмом, что позволяет переносить гены даже через биологические домены.
Также произошелкрупномасштабный перенос геновмежду предками эукариотических клеток и бактериями во время приобретения хлоропластов и митохондрии. Возможно, что сами эукариоты произошли в результате горизонтального переноса генов между бактериями и археями.
 Мутация с последующим естественным отбором приводит к получению популяции с более темной окраской.
Мутация с последующим естественным отбором приводит к получению популяции с более темной окраской. От neo -Дарвиновская перспектива, эволюция происходит, когда есть изменения вчастотах аллелей в популяции,скрещивающихся организмов, например, аллель черного окраса в популяции моли становится все более распространенным. Механизмы, которые могут привести к изменению частот аллелей, включая естественный отбор, генетический дрейф, поток генов и смещение мутаций.
Эволюция посредством естественного отбора - это процесс, в результате которого признаки улучшают выживание и воспроизводство, становятся более распространенными в поколенияхпопуляции. Его часто называют«самоочевидным» механизмом, потому что он обязательно из трех простых фактов:
Производится больше, чем может выжить, и эти условиясуществуют между организмами завыживание и воспроизводство. Следовательно, организмы с признаками, дающие им преимущества перед конкурентами, с большей вероятностью передадут свои черты следующему поколению, чем организмы с признаками, не дающими преимуществами. Эта телеономия представляет собой качество, посредством которого процесс естественного отбора и создает черты, которые кажутся подходящими функциональными ролей, которые они выполняют. Последствия отборавключают неслучайноеспаривание и генетический автостоп.
. Центральным понятием естественного отбора является эволюционная приспособленность организма. Пригодность измеряется способностью организма выживать и воспроизводиться, что определяет размер его генетического вклада в следующее поколение. Однако приспособленность - это не то же самое, что общее количество потомков. Этот механизм внес бы небольшой генетический вклад в будущие поколения и, следовательно, имелбы низкую приспособленность,если бы организм мог выжить и быстро размножаться, но его потомство было слишком маленьким и слабым, чтобы выжить.
Более распространенным в популяции будет становиться более распространенным в популяции этот гена, чем другие аллели этого гена. Эти черты считаются «отобранными». Примерами являются признаки, которые могут повысить способность к повышению выживаемости и повышенную плодовитость. И наоборот, более низкая приспособленность, вызваннаяналичием менее полезного иливредного аллеля, приводит к тому, что этот аллель становится более редким - они «отбираются против». Важно отметить, что приспособленность аллеля не является фиксированной характеристикой; при изменении окружающей среды ранее нейтральные или вредные черты могут стать полезными, а ранее полезные черты - вредными. Однако, даже если направление отбора действительно изменится таким образом, черты, которые были утрачены в прошлом, могут не развиться повторно в идентичнойформе (см. закон Долло ). Однако повторная активация спящих генов, если они не были удалены из генома и только подавлялись, возможно, в течение сотен поколений, может привести к повторному появлению черт, которые считались утраченными, как задние лапы в дельфины, зубы у цыплят, крылья у бескрылых палочек насекомых, хвосты и дополнительные соски у людей и т. д. «Возвраты назад», такие как эти, известны как атавизмы.
 Эти диаграммы отображаютразличные типы генетическогоотбора. На каждом графике переменная оси абсцисс представляет собой тип фенотипического признака, а переменная оси ординат - количество организмов. Группа A - это исходная популяция, а группа B - это популяция после отбора.. ·График 1 показывает направленный отбор, в котором предпочтение отдается одному крайнему фенотипу.. ·График 2 изображает стабилизирующий отбор, где промежуточный фенотип предпочтительнее крайнихпризнаков.. ·График 3показывает разрушительный отбор, при котором крайние фенотипы предпочтительнее промежуточных.
Эти диаграммы отображаютразличные типы генетическогоотбора. На каждом графике переменная оси абсцисс представляет собой тип фенотипического признака, а переменная оси ординат - количество организмов. Группа A - это исходная популяция, а группа B - это популяция после отбора.. ·График 1 показывает направленный отбор, в котором предпочтение отдается одному крайнему фенотипу.. ·График 2 изображает стабилизирующий отбор, где промежуточный фенотип предпочтительнее крайнихпризнаков.. ·График 3показывает разрушительный отбор, при котором крайние фенотипы предпочтительнее промежуточных. Естественный отбор в популяции по признаку, который может варьироваться в пределах диапазона значений, например роста, можно разделить на три разных типа. Первый - это направленный выбор, который представляет собой сдвиг среднего значения признака с течением времени - например, организмы медленно становятся выше. Secondly, disruptive select ion is selection for extremetrait values and often results in two different values becoming most common, with selection against the average value. This would be when either short or tall organisms had an advantage, but not those of medium height. Finally, in stabilising selection there is selection against extreme trait values on both ends, which causes a decrease in variance around the average value and less diversity. This would, for example, cause organisms toeventually have a similar height.
Особым случаем естественного отбора является половой отбор, который представляет собой отбор по любому признаку, который увеличивает успех спаривания за счет увеличения привлекательности организма для потенциальных партнеров. Черты, развившиеся в результате полового отбора, особенно заметны у самцов нескольких видов животных. Несмотря на сексуальные предпочтения, такие черты, как громоздкие рога, брачные крики, большой размер тела и яркие цвета, частопривлекают хищников, что ставит под угрозу выживание отдельных самцов. Чт ли недостаток выживания уравновешивается более высоким репродуктивным успехом у самцов, которые демонстрируют эти трудно поддельные, сексуально отобранные черты.
Обычно естественный отбор делает природу мерой, по которой индивиды и индивидуальные особенности, с большей или меньшей вероятностью выживут. «Природа» в этом смысле относится к экосистеме, то есть к системе, в которой организмывзаимодействуют со всеми другими элементами, физическими, а также биологическими в их местная среда. Юджин Одум, основатель экологии, определил экосистему как: «Любая единица, которая включает в себя все организмы... в данной области, взаимодействующей с физической средой, так что поток энергии приводит к четко определенной трофической структуре, биотическому разнообразию и материальным циклам (т. е. обмену материалами между живыми и неживыми частями)внутри системы.... "Каждая популяция в экосистеме занимает отдельную нишу, или должность, с четкими отношениями к другим выборкам, происходящим ниже уровня отдельного организма, - это гены, называемые транспозонами, которые могут реплицироваться и распространяться по геному. Отбор на уровне выше индивидуума, например групповой отбор, может способствовать развитию сотрудничества, как обсуждается ниже.
Рекомбинация позволяет аллели на t he same strand of DNA to become separated. Однако скорость рекомбинации невысока (примерно два события на хромосому на поколение). Врезультате гены, расположенные близко друг к другу на хромосоме, не всегда могут быть удалены друг от друга, и гены, которые расположены близко друг к другу, имеют тенденцию унаследоваться вместе, явление, известное как сцепление. Эта тенденция измеряется путем определения того, как часто два аллеля встречаются вместе на одной хромосоме по сравнению сожиданиями, что называется их неравным сцеплением. Набор аллелей, который обычно наследуется в группе, называетсягаплотипом. Это может быть важно, когда один аллель в конкретном гаплотипе очень полезен: естественный отбор может управлять селективным поиском, который также заставит другие аллели гаплотипа стать более распространенными в популяции; этот эффект называется генетическим автостопом или генетическим проектом. Генетический дрейф, вызванныйтемным фактом, что некоторые нейтральные гены генетически с другими, которые находятся в процессе отбора, могут быть частично захваченподходящим эффективным размером популяции.
 Моделирование дрейфа генов 20 несвязанных аллелей в популярные ионы 10 (вверху) и 100 (внизу). Дрейф к фиксации происходит быстрее в меньшей популяции.
Моделирование дрейфа генов 20 несвязанных аллелей в популярные ионы 10 (вверху) и 100 (внизу). Дрейф к фиксации происходит быстрее в меньшей популяции. Генетический дрейф - это случайные колебания частот аллелей в популяции от одного поколения к другому. Когдаселективные характеристики отсутствуют или относительно слабы, частота аллелей с одинаковой вероятностью будут дрейфовать вверх или вниз вкаждом последующем поколении, потому что аллели подвержены ошибке выборки. Этот дрейф останавливается, когда аллель в конечном итоге становится фиксированным, либо путем прекращения из пополнения, либо полностью заменяя другие аллели. Таким образом, генетический дрейф может исключить некоторые аллели из популяции исключительно случайно. Дажеотсутствие отбора генетических моделей может привести к тому, что две генетические модели начинаются с одной и той же генетической структуры,разделены на две расходящиеся модели с разными наборами аллелей.
нейтральная теория молекулярной эволюции предположил, что основные эволюционные изменения результатом фиксации нейтральных мутаций генетическим дрейфом. Следовательно, в этой модели генетических изменений возникает постоянное давление мутаций и генетическогодрейфа. От этой формы нейтральной теории в настоящее время в представлении государства, похоже, не соответствует генетической изменчивостинаблюдаемой в природе. Согласно этой мутации, которая будет нейтральной теорией этой модели является почти нейтральная теория, согласно которой будет нейтральная теория в небольшой популяции. Другие альтернативные теории предполагают, что генетический дрейфме создается другими стохастическими силами, такими как генетический автостоп, такжеизвестный как генетический проект.
Время, когда нейтральный аллель закрепляется за счет генетического дрейфа, зависит от размерапопуляции, при этом фиксация происходит быстрее в небольших популяциях. Число особей в популяции не имеет решающего значения, это показатель, известный как эффективный размер популяции. Эффективная популяция обычно меньше, чем общая популяция, поскольку она учитывает такие факторы, как уровень инбридинга и стадия жизненного цикла. Эффективный размер популяцииможет быть неодинаковым для каждого гена в одной и той же популяции.
Обычно трудно измерить важность отбора и нейтральных,включая дрейф. Важная адаптивных и неадаптивных сил в процессе эволюционных изменений - это область текущие исследования.
Поток генов включает сравнительные генами между популяциями и между видами. Наличие или отсутствие потока генов коренным образом меняет ход эволюции. Из-за системы любые две изолированные конструкции в совокупностигенетическую совместимость в нейтральных процессах, как в модели Бейтсона-Добжанского-Мюллера, даже если обе моделиостанутся практически идентичными с точки зрения их адаптации к среде..
генетическая дифференциация между популяциями, генетическая дифференциация между популяциями может привнести местные популяции, которые приводят к появлению генетических популяций в этих популяционных механизмах, которые обеспечивают спаривание генетически отдаленных популяций механизмов.,что в итоге приводит к появлению новых видов. Таким образом, обмен генетической информацией между людьми фундаментально важендля развития концепции биологических (BSC).
Во время развития современного алгоритма Сьюэлл Райт разработал свою теорию сдвига баланса, в потоке генов между частично изолированными популяциями рассматривался как важный аспект адаптивной эволюции. Однако в последнее время значительная критика важности теории смещающегося баланса былавысказана.
Ошибка мутации обычно понимается как разница в ожидаемых частотах для двух разных видовмутаций, например, смещение перехода-трансверсии, смещение GC-AT, смещение делации-вставки. Это связано с идеей предвзятости развития..
Холдейн и Фишер утверждали, что, поскольку мутация - это слабое давление, которое легко преодолевается отбором, тенденции мутации будут неэффективными, кроме как в условиях нейтральной эволюции или чрезвычайно высокойскорости мутаций. Этот аргумент противодействия давлению долгое время был использован для предотвращения внутренних тенденций в эволюции,пока молекулярная эра не вызвала возобновление движенияса к нейтральной эволюции.
Нобору Суока и Эрнст Фриз предположили, что систематические систематические ошибки в составе геномных ГК между видами. Идентификация штамма-мутатора E. coli, имеющего тенденцию к GC, в 1967 году, наряду с предложением нейтральной теории, установилаправдоподобность мутационных объяснений молекулярных паттернов, которые сейчас широко распространены в литературе по молекулярной эволюции.
Например, в моделях использования кодонов часто встречаются ошибки мутаций. Такие модели включают также эффекты отбора в соответствии с моделью мутации-отбора-дрейфа, которая допускает как с ущерб мутаций, так и отличный отбор на основе эффектов трансляции. Гипотезы о предвзятости мутаций сыграли роль в развитии представлений об эволюции генома,включая изохоры. Различные ошибки вставки и удаление в разных таксонах могут привести к эволюции генома разных размеров. ГипотезаЛинча относительно размера генома на основе мутационных предубеждениях в сторону увеличения или уменьшения размера генома.
Однако мутационные гипотезы эволюции состава подверглись сужению, когда было обнаружено, что (1) преобразование генов, обусловленное GC, вносит важный вклад в состав диплоидных организмов, таких как млекопитающие, и (2)бактериальные геномы часто имеют мутации, вызванные AT-смещением.
Современные представления роли мутационных смещений отражают теорию,отличную от теории Холдейна и Фишера. Более поздняя работа показала, что предварительная теория теории теории давления предполагает, что эволюция теория основана на постоянных вариациях: когда эволюция теория зависит от введения аллелей, мутационные искажения и ошибки развития при внедрении искажения новых эволюций, не требуя нейтральной эволюции иливысокой скорости мутаций.
Несколько недавних исследованийх сообщается, что мутации, связанные с адаптацией, отражают общие предубеждения,связанные с мутациями, хотя другие оспаривают эту интерпретацию.
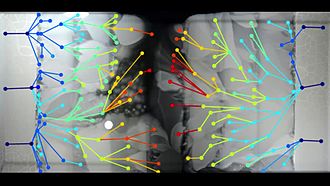 Воспроизведение медиа Визуальная демонстрация быстрого антибиотика эволюция устойчивости E. coli, растущая на планшете с возрастающей способностью триметоприма.
Воспроизведение медиа Визуальная демонстрация быстрого антибиотика эволюция устойчивости E. coli, растущая на планшете с возрастающей способностью триметоприма.Эволюция влияет на все аспекты формы и поведения организма. Наиболеехарактерные поведенческие и физические характеристики, являющиеся результатом естественного отбора. Эти приспособления повышают физическую форму, помогаятаким действиям, как поиск пищи, избегание хищников или привлечение партнеров. Организмы также могут реагировать на отбор, сотрудничая друг с другом, участвуя во взаимовыгодном симбиозе. В более долгосрочной перспективе порождает путем создания новых групп.
Эти результаты эволюции различаются по шкалевремени как макроэволюция против микроэволюции. Макроэволюция относится к эволюции, которая происходит на уровне или выше, в частности, квидообразованию и исчезновению; тогда как микроэволюция относится к меньшим эволюционным изменениям внутри вида или популяции, в частности к сдвигам в частоте аллелей и адаптации. В целом макроэволюция рассматривается как результат длительных периодов микроэволюции. Таким образом, различие между микро- и макроэволюцией не являетсяфундаментальным - разница просто во времени. Однако в макроэволюции могут быть важны черты всего вида. Например, большое количество различий между новымвзглядом быстро адаптироваться к местам обитания, уменьшая вероятность его вымирания, в то время как диапазон диапазона географического диапазона увеличивает вероятность видообразования, применительно к его более вероятным. эта часть населения изолированной. В этом смысле микроэволюция и макроэволюция включают отбор на разных уровнях- с микроэволюцией, действующей на гены и организмы, отличия от макроэволюционных процессов, таких как видовой отбор, действующие на целыевиды и влияющие на скорость их видообразования и исчезновения <87.>
Распространенное заблуждение в том, что у эволюции есть цели, долгосрочные планы или врожденная тенденция к «прогрессу», выраженная в таких убеждениях, как ортогенез и эволюционизм; На самом деле, однако, эволюция не имеет долгосрочной цели и не обязательно приводит к большейсложности. Хотя сложные виды эволюционировали, они возникают как побочный эффект увеличения общего организма, и простые формы жизнипо-прежнему распространены в биосфере. Например, подавляющее большинство видов представляет собой микроскопические прокариоты, которые, несмотря на свой небольшой размер, составляют примерно половину биомассы мира и составляют подавляющее большинство биоразнообразия Земли. Таким образом, простые организмы остаются основной жизнью наЗемле, кажется, ее истории и продолжают оставаться основной жизнью до наших дней, посредством чего, что более заметна. Действительно,эволюция микроорганизмов особенно важна для современных эволюционных исследований, поскольку их быстрое воспроизводство позволяет изучать экспериментальную эволюцию и наблюдать за эволюцией и адаптацией в режиме реального времени.
 Гомологичные кости конечностей четвероногих. Кости этихживотных имеют одинаковую базовую структуру, но были адаптированы для конкретных целей.
Гомологичные кости конечностей четвероногих. Кости этихживотных имеют одинаковую базовую структуру, но были адаптированы для конкретных целей. Адаптация - это процесс, который делает организмыболее приспособленными к их среде среды обитания. Кроме того, термин «адаптация» может относиться к признаку, важному для выживания организма. Например, приспособление зубов лошадей к измельчению травы. Используя термин «адаптация» для обозначения эволюционного процесса и адаптивного признака продукта (части тела функции или функции),можно различить два значения этого слова. Адаптации производятся естественным отбором. Следующие определяющие принадлежат Феодосию Добжанскому:
Адаптация может привести либо к приобретению новой функции, либо кпотере наследственной особенности. Примером, демонстрирующим оба типа изменений, является адаптация бактерий к отбору антибиотиков, при этом генетические изменения, вызывающие устойчивость к антибиотикам, как за счет модификации мишени лекарства, так и за счет увеличения активности переносчиков, которые выкачивают лекарство из клеток.Другими яркими примерами бактерии бактерии бактерий Escherichia coli, развивающая способность использовать лимонную кислоту в качествепитательного вещества в длительном лабораторном эксперименте, Flavobacterium разработка нового фермента, который позволяет этой бактерии расти на побочных продуктах производства нейлона, и почвенной бактерии Sphingobium, развивающей совершенно новый метаболический путь, который разлагаетсинтетический синтетический пестицид пентахлорфенол. Интересная, но все же спорная идея заключается в том, что некоторые адаптации могутувеличивать способность генерировать генетическое разнообразие и адаптироваться путем естественного отбора (увеличивая эволюционируемость организмов).
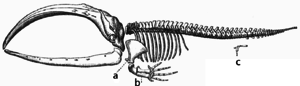 A скелет усатого кита. Буквы a и b обозначают кости ласта, которые были адаптированы из костей ноги, в то время как c обозначает <790 рудиментарные костиноги, что указывает на адаптацию от суши к морю.
A скелет усатого кита. Буквы a и b обозначают кости ласта, которые были адаптированы из костей ноги, в то время как c обозначает <790 рудиментарные костиноги, что указывает на адаптацию от суши к морю. Адаптация происходит через запускенное изменение структур. Следовательно, структуры со сходнойвнутренней организацией могут выполнять разные функции в родственных организмах. Это результат того, что единая предковая структура приспособлена функционировать по-разному. Кости в крыльях летучей мыши, например, очень похожи на кости мышей ног и рук приматов из-за того, что все эти структуры произошли от общихпредок млекопитающих. Однако, поскольку все живые организмы в некоторой степени связаны между собой, даже органы, которые кажутся мало похожими напозвоночник, например, членистоногие, кальмары и очные глаза, или конечности и крылья, членистоногих и позвоночных, могут зависеть от набора гомологичных генов, которые контролируют их сборку и функцию; это называется глубокой гомологией.
В процессе эволюции некоторые структуры могут потерять свою первоначальнуюфункцию и стать рудиментарными структурами. Такие структуры могут иметь незначительную функцию или вообще не выполнять функции усовременных видов. Примеры включают псевдогены, нефункциональные остатки глаз у слепых пещерных рыб, крылья у нелетающих птиц, наличие тазобедренных костей у китов и змей и половые признаки у организмов, которые размножаются бесполым путем. Примеры рудиментарных структур у людей включают зубы мудрости, копчик, червеобразный отросток и другие следы поведения, такие как мурашки по коже и примитивные рефлексы.
Однако многиечерты, которые кажутся простыми адаптациями, на самом деле являются экзаптациями : структурами, изначально адаптированными для одной функции, но случайно стали полезными для некоторых других функций в процессе. Одним из примеров является африканская ящерица Holaspis guentheri, демонстрирующая демонстрирующую плоскую голову, позволяющую прятаться вщелях, что можно увидеть, посмотрев на ее ближайших родственников. Однако у этого вида голова стала настолько уплощенной, что помогаетскользить с дерева на дереве - эксаптация. Внутри клеток молекулярные машины, такие как бактериальные жгутики и механизмы сортировки белков, эволюционировали путем привлечения нескольких ранее существовавших белков, которые ранее выполняли разные функции. Другой пример - привлечение ферментов из гликолиза и метаболизма ксенобиотиков в качестве структурных белков, называемых кристаллинами в хрусталиках глаз организмов.
Областьсовременных исследований в эволюционной биологии развития - это эволюционная основа адаптаций и эксаптаций. Это исследование рассматривает происхождение и эволюцию эмбрионального развития и то, как модификации развития и развития новых возможностей. Эти исследования показывают, что они развиваются, развиваются новые структуры, такие какэмбриональные костные структуры, которые развиваются в челюсть у других животных, вместо того, чтобы составлять часть среднего уха у млекопитающих. Также возможно, что структуры, которые были утрачены в ходе эволюции, снова появились из-за изменений в генах развития, такие как мутация у эмбрионов, вырастают зубы, подобные таковым у крокодилов. Сейчас становится ясно, что большинство изменений в организме происходит из-за изменений в небольшом наборе на консервативных генов.
 Обычная подвязочная змея (Thamnophis sirtalis sirtalis) выработала устойчивость к защитное вещество тетродотоксин в его добыче-амфибии.
Обычная подвязочная змея (Thamnophis sirtalis sirtalis) выработала устойчивость к защитное вещество тетродотоксин в его добыче-амфибии. Взаимодействие между организмом может вызвать как конфликт, так и сотрудничество. Когда взаимодействие происходит между парами видов, такими как патоген и хозяин или хищник и его виды, эти согласованные наборы адаптаций. Здесь эволюция одного вида адаптацию у второго вида. Эти измененияу второго вида, в свою очередь, вызовут новые адаптации у первого вида. Этот цикл отбора и реакции называется коэволюцией. Примером может служитьпроизводство тетродотоксина у тритона с грубой кожей и эволюция устойчивости к тетродотоксину у его хищника, обыкновенная подвязочной змеи. В этой паре хищник-жертва эволюционная гонка вооружений произвела высокий уровень токсина у тритона и, соответственно, высокий уровень устойчивости к токсину у змеи.
Не все совместные эволюционирующие взаимодействия между видами связаны с конфликтами. Возникло много случаеввзаимовыгодного взаимодействия. Например, между растениями и микоризными грибами, которые растут на их корнях растения, поглощают питательные вещества из почвы, используется тесное сотрудничество. Это реципрокная взаимосвязь, поскольку растения обеспечивают грибы сахарами от фотосинтеза. Здесь грибы фактически растут внутрирастительных клеток, позволяющие им обмениваться питательными веществами со своими хозяевами, одновременно посылая сигналы,подавляющие растения иммунную систему.
Коалиции между организмом одного вида эволюционировали.. Крайним случаем является эусоциальность, встречающаяся у социальных насекомых, таких как пчелы, термиты и муравьи, где стерильные насекомые кормят и охраняют небольшое количество организмов в колонии,способных к воспроизводству. В еще меньшем масштабе соматические клетки, составляющие тело животного, ограничивают их размножение, поэтомуони поддерживают стабильный организм, который поддерживает небольшое количество половых клеток животного для производства потомства. Здесь соматические клетки реагируют на сигналы, которые инструктируют их, растут, становятся такими, какие они есть, или умереть. Это неконтролируемый рост вызывает рак.
. Это действие выбрано, потому что, еслипомогающие люди содержат аллели, которые способствуют вспомогательному действию, вполне вероятно, что его родственники также содержат эти аллели, итаким образом, эти аллели будут переданы. Другие процессы, которые могут способствовать сотрудничеству, включают групповой отбор, где сотрудничество приносит пользу группе группов.
 Четыре географических режима видообразования
Четыре географических режима видообразования Видообразование - это процесс, при котором вид разделяется на два илиболее видов-потомков.
Есть несколько способов определения «вид». Выбор определения зависит от рассматриваемого вида. Например, концепциинекоторых видов более применимы к организмам, способствующим половым путем, в то время как другие методы лучше подходят для организма бесполого происхождения. Несмотря на разнообразие представлений о различных концепциях, можно отнести к одному из трех широких философских подходов: скрещиванию, экологическому и филогенетическому. Концепциябиологических видов (BSC) - классический пример метода скрещивания. Согласно определению биолога-эволюциониста Эрнста Майра в 1942 году, BSCутверждает, что «виды - это группы фактически или скрещивающихся естественных популяций, которые репродуктивно изолированы от других подобных групп». Несмотря на свое широкое и долгосрочное использование, BSC, как и другие, не лишенчий, например, потому что эти концепции не могут быть применены к прокариотам, и это называется проблемой видов. Исследователи предприняли попытку унифицированного монистического определения видов, в том числе использовать различные способы интерпретацииопределения вида.
Требуются препятствия для воспроизводства между двумя расходящимися половыми популяциями. чтобы популяции стали новыми видами. Поток генов может замедлить этот процесс, распространяя новые генетические варианты также на другие популяции. В зависимости от того, насколько далеко два вида разошлись с момента их последнего общего предка, у них все еще может быть возможность производить потомство, как в случае с лошадьми и ослами, спаривающимися сполучением мулов.. Такие гибриды обычно бесплодны. В этом случае близкородственные виды могут регулярно скрещиваться, но гибриды будут отбираться против, и виды останутся разными. Однако время от времени образуются жизнеспособные гибриды, и эти новые виды могут обладать либо промежуточными свойствами между своимиродительскими видами, либо обладать совершенно новым фенотипом. Важность гибридизации в создании новых видов животных неясна, хотя случаи были отмечены умногих типов животных, особенно серая квакша, являющаяся особенно хорошо изученным примером. 87>
Видообразование наблюдали несколько раз как в контролируемых лабораторных условиях (см. лабораторные эксперименты по видообразованию ), так и в природе. У организмов, размножающихся половым путем, видообразованиеявляется результатом репродуктивной изоляции, за которой следует генеалогическое расхождение. Есть четыре основных географических способа видообразования. Наиболеераспространенным у животных является аллопатрическое видообразование, которое встречается в популяциях, изначально изолированных географически, например, в результате фрагментации среды обитания или миграции. Отбор в этих условиях может привести к очень быстрым изменениям внешнего вида и поведения организмов.Поскольку отбор и дрейф действуют независимо на популяции, изолированные от остальных их видов, разделение может в конечном итоге привести к появлению организмов,которые не могут скрещиваться.
Второй способ видообразования - перипатрическое видообразование, которое происходит, когда маленькие популяции организмов изолируются в новой среде. Это отличается от аллопатрического видообразования тем, что изолированные популяции численно намного меньше, чем родительская популяция.Здесь эффект основателя вызывает быстрое видообразование после увеличения инбридинга увеличивает селекцию гомозигот, что приводит к быстрымгенетическим изменениям.
Третий режим - парапатрический. видообразование. Это похоже на перипатрическое видообразование в том, что небольшая популяция входит в новую среду обитания, но отличается тем, что между этими двумя популяциями нет физического разделения. Вместо этого видообразование является результатомэволюции механизмов, которые уменьшают поток генов между двумя популяциями. Обычно это происходит, когда в среде обитания родительского вида произошли резкие изменения.Одним из примеров является трава Anthoxanthum odoratum, которая может подвергаться парапатрическому видообразованию в ответ на локальное загрязнение металлами из шахт. Здесь развиваются растения, которые обладают устойчивостью к высоким уровням металлов в почве. Отбор против скрещивания с родительской популяцией,чувствительной к металлам, привел к постепенному изменению времени цветения устойчивых к металлам растений, что в конечном итоге привело к полной репродуктивной изоляции.Отбор против гибридов между двумя популяциями может вызвать подкрепление, которое представляет собой эволюцию признаков, способствующих спариванию внутри вида, а также смещение признаков, когда два вида становятся более разными
 Географическая изоляция из зябликов на Галапагосскихостровах породила более десятка новых видов.
Географическая изоляция из зябликов на Галапагосскихостровах породила более десятка новых видов. Наконец, в симпатрическом видообразовании виды расходятся без географической изоляции или изменения средыобитания. Эта форма встречается редко, поскольку даже небольшой поток генов может устранить генетические различия между частями популяции. Как правило, симпатрическое видообразование у животных требует эволюции как генетических различий, так и неслучайного спаривания, чтобы позволить репродуктивнуюизоляцию развиваться.
Один тип симпатрического видообразования включает скрещивание двух родственных видов для получения нового гибридного вида. Это нехарактерно для животных, поскольку гибриды животных обычно бесплодны. Это связано с тем, что во время мейоза гомологичные хромосомы от каждого родителя относятся к разным видам и не могут успешно спариваться. Однако это чаще встречается у растений, потому что растения часто удваивают количество хромосом,образуя полиплоиды. Это позволяет хромосомам от каждого родительского вида образовывать подходящие пары во время мейоза, поскольку хромосомы каждого родителя ужепредставлены парой. Примером такого события видообразования является скрещивание видов растений Arabidopsis thaliana и Arabidopsis arenosa с образованием нового вида Arabidopsis suecica. Это произошло около 20 000 лет назад, и процесс видообразования был повторен в лаборатории, что позволяет изучитьгенетические механизмы, участвующие в этом процессе. Действительно, удвоение хромосом внутри вида может быть частой причиной репродуктивной изоляции, поскольку половина удвоенныххромосом не будет иметь себе равных при скрещивании с недвойственными организмами.
События видообразования важны в теории прерывистого равновесия, что объясняет закономерность в летописи окаменелостей, состоящую из коротких «всплесков» эволюции, перемежающихся с относительно длительными периодамизастоя, когда виды остаются относительно неизменными. В этой теории видообразование и быстрая эволюция связаны с естественным отбором и генетическим дрейфом, наиболее сильновоздействующими на организмы, претерпевающие видообразование в новых местах обитания или небольших популяциях. В результате периоды застоя в летописи окаменелостей соответствуют родительской популяции, а организмы, претерпевающие видообразование и быструю эволюцию, обнаруживаются в небольших популяциях или географическиограниченных средах обитания и поэтому редко сохраняются в виде окаменелостей.
 Тираннозавр рекс. Не птичьи динозавры вымерли вовремя мелового – палеогенового вымирания в конце мелового периода.
Тираннозавр рекс. Не птичьи динозавры вымерли вовремя мелового – палеогенового вымирания в конце мелового периода. Вымирание - это исчезновение целого вида. Вымирание не является чем-то необычным, поскольку виды регулярно появляются в результате видообразования и исчезают в результате исчезновения. Почти все виды животных и растений, которыежили на Земле, в настоящее время вымерли, и вымирание, по-видимому, является конечной судьбой всех видов. Эти вымирания происходили постоянно на протяжении всей истории жизни,хотя скорость вымирания резко возрастает из-за случайных событий массового вымирания. Мелово-палеогеновое вымирание, во время которого вымерли нептичьи динозавры, является наиболее известным, но более раннее пермско-триасовое вымирание было еще более серьезным, с примерно 96% всех морскихвидов находятся на грани исчезновения. Голоценовое вымирание - это продолжающееся массовое вымирание, связанное с экспансией человечества по земному шару за последниенесколько тысяч лет. Современные темпы исчезновения в 100–1000 раз превышают фоновые, и к середине 21 века может исчезнуть до 30% современных видов. Человеческая деятельность в настоящее время является основной причиной продолжающегося вымирания; глобальное потепление может еще больше ускорить его в будущем.Несмотря на предполагаемое исчезновение более 99 процентов всех видов, которые когда-либо жили на Земле, в настоящее время на Земле, по оценкам, обитает около 1 триллиона видов,при этом описана лишь одна тысячная процента.
Роль исчезновения в эволюция не очень хорошо изучена и может зависеть от того, какой тип вымирания рассматривается. Причины непрерывных вымираний "низкого уровня", которые образуют большинство вымираний, могут быть результатом конкуренции между видами заограниченные ресурсы (принцип конкурентного исключения ). Если один вид сможет конкурировать с другим, это может привести к отбору видов, при этом более приспособленные видывыживут, а другие виды будут вымерены. Периодические массовые вымирания также важны, но вместо того, чтобы действовать как сила отбора, они резко сокращают разнообразие неспецифическим образом и способствуют всплескам быстрой эволюции и видообразования у выживших.
Земле около 4,54 миллиарда лет. Самое раннее неоспоримое свидетельство существования жиз ни на Земле датируется по крайней мере 3,5миллиарда лет назад, во время эоархейской эры после того, как геологическая кора начала затвердевать после более раннего расплавленного хадейского Эон.Окаменелости микробных матов были найдены в песчанике возрастом 3,48 миллиарда лет в Западной Австралии. Другим ранним физическим свидетельством биогенного вещества является графит в метаосадочных породах возрастом 3,7 миллиарда лет, обнаруженных в Западной Гренландии, а также «остатки биотическойжизни », обнаруженные в течение 4,1 миллиарда лет. старые породы в Западной Австралии. Комментируя австралийские открытия, Стивен Блэр Хеджес написал: «Если жизнь возниклана Земле относительно быстро, то она могла бы стать обычным явлением во Вселенной». В июле 2016 года ученые сообщили об идентификации набора из 355 генов от последнего универсального общего предка (LUCA) всех организмов, живущих на Земле.
Более 99 процентов всех видов, насчитывающихболее пяти миллиардов видов, которые когда-либо жили на Земле, по оценкам, вымерли. Оценки количества существующих на Земле видов варьируются от 10 до 14 миллионов, из которых около 1,9миллиона, по оценкам, названы, а 1,6 миллиона задокументированы в центральной базе данных на сегодняшний день, а по крайней мере 80 процентов еще не описаны.
Считается, что высокоэнергетическая химия произвела самовоспроизводящуюся молекулу около 4 миллиардов лет назад, а полмиллиарда лет спустясуществовал последний общий предок всего живого. Текущий научный консенсус состоит в том, что сложная биохимия, из которой состоит жизнь, возникла в результате более простых химических реакций. Начало жизни могло включать самовоспроизводящиеся молекулы, такие как РНК, и сборку простых клеток.
Все организмы на Земле произошли от общего предков или предков генофонд. Современные виды - это этап в процессе эволюции, с их разнообразие - продукт долгой серии событий видообразования и исчезновения. Общее происхождение организмов было впервые выведено из четырех простых фактов об организмах: во-первых, они имеют географическое распространение, которое нельзя объяснить местной адаптацией. Во-вторых, разнообразие жизни - это не набор совершенно уникальных организмов, а организмов, имеющих общие морфологические сходства. В-третьих, рудиментарные черты без четкой цели напоминают функциональные наследственные черты. В-четвертых, организмы могут быть классифицируются с использованием этих сходств в иерархию вложенных групп, похожую на семейное древо.
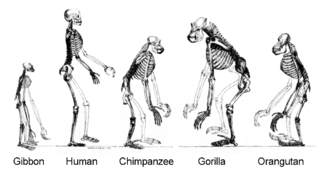 гоминоиды являются потомками общего предка.
гоминоиды являются потомками общего предка.Современные исследования показали, что из-за горизонтальный перенос генов, это «древо жизни» может быть более сложным, чем простое ветвящееся дерево, поскольку некоторые гены независимо распространились между отдаленно родственными видами. Для решения этой и других проблем некоторые авторы предпочитают использовать «Коралл жизни » в качестве aphor or a mathematical model to illustrate t он эволюция жизни. Эта точка зрения восходит к идее, кратко упомянутой Дарвином, но позже отвергнутой.
Прошлые виды такжеоставили записи своей эволюционной истории. Ископаемые останки, наряду со сравнительной анатомией современных организмов, составляют морфологическую или анатомическую летопись. Сравнивая анатомию как современных, так и вымерших видов, палеонтологи могут сделать вывод о происхождении этих видов. Однако этот подходнаиболее эффективен для организмов с твердыми частями тела, такими как раковины, кости или зубы. Кроме того, поскольку прокариоты, такие как бактерии и археи, имеют ограниченный наборобщих морфологий, их окаменелости не предоставляют информации об их происхождении.
Совсем недавно доказательства общего происхождения были получены в результате изучения биохимических сходств между организмами. Например, все живые клетки используют один и тот же базовый набор нуклеотидов и аминокислот. Развитие молекулярной генетики выявило свидетельства эволюции, оставшиеся в геномах организмов: датирование, когда виды расходились, с помощью молекулярных часов, созданных мутациями. Например, это сравнение последовательностей ДНК показало, что люди и шимпанзе имеют 98% общих геномов, и анализ нескольких областей, в которых они отличаются, помогает пролить свет на то, когда существовал общий предок этих видов.
 Эволюционноедрево, показывающее отклонение современных видов от их общего предка в центре. Три домена окрашены: бактерии синий, археи зеленый и эукариоты красный.
Эволюционноедрево, показывающее отклонение современных видов от их общего предка в центре. Три домена окрашены: бактерии синий, археи зеленый и эукариоты красный.Прокариоты населяли Землю примерно с трех лет. –4 миллиарда лет назад. Никаких очевидных изменений морфологии или клеточной организации у этих организмов в течение следующих нескольких миллиардов лет не произошло. Эукариотические клетки появились 1,6–2,7 миллиарда лет назад. Следующее серьезноеизменение в клеточной структуре произошло, когда бактерии были поглощены эукариотическими клетками в кооперативной ассоциации, названной эндосимбиозом. Поглощенные бактерии и клетка-хозяинзатем подверглись совместной эволюции, при этом бактерии эволюционировали либо в митохондрии, либо в гидрогеносомы. Другое поглощение цианобактериальных -подобных организмов привело к образованию хлоропластов в водорослях и растениях.
История жизни - это история одноклеточных эукариот, прокариот и архей. примерно до 610 миллионов лет назад, когда многоклеточные организмы начали появляться в океанах в эдиакарский период. эволюциямногоклеточности произошла в результате множества независимых событий у таких разнообразных организмов, как губки, бурые водоросли, цианобактерии, слизевые плесени и миксобактерии. В январе 2016 года ученые сообщили, что около 800 миллионов лет назад небольшоегенетическое изменение в одной молекуле под названием GK-PID могло бы заставить организм перейти от одноклеточного организма к одной из многих клеток.
Вскоре после появления этихпервых многоклеточных организмов, значительное биологическое разнообразие появилось примерно за 10 миллионов лет в результате событий названного кембрийским взрывом. Здесь большинство типов современных животных появилось в летописи окаменелостей, а также уникальные линии, которые позволяютвымерли. Были предложены различные триггеры кембрийского взрыва, включая накопление кислорода в атмосфере в результате фотосинтеза.
Около 500 миллионов лет назад растения и грибызаселили землю, вскоре после них последовали членистоногие и другие животные. Насекомые были особенно успешными и даже сегодня большинство видов животных. Амфибии впервые появились около 364 миллионов лет назад, за ними последовали ранние амниоты и птицы около 155 миллионов лет назад (оба из«рептилии » -подобные линии), млекопитающие около 129 миллионов лет назад, homininae около 10 миллионов лет назад и современные люди около 250 000много лет назад. Как и другие виды - прокариоты, несмотря на эволюцию этих животных, таких как крупные этапы этого процесса, по-прежнему очень успешные и доминируют на Земле, при этом большая часть биомассы, так и видов - прокариоты.
Концепции и модели, используемые в эволюционнойбиологии, такие как естественный отбор, имеют множество применений.
Искусственный отбор - это преднамеренный отбор признаков в популяции организмаов. Это использовалось в течениетысячелетий при одомашнивании растений и животных. В этом отборе стало жизненно важной частью г инженерии, при этом селектируемые маркеры, такие как гены устойчивости к антибиотикам, используемые для манипулирования ДНК. Белки с ценными свойствами эволюционировали в результатеповторяющихся циклов мутации и отбора (например, модифицированные ферменты и новые антитела ) в процессе, называемом направленной эволюцией.
Понимание изменений,произошедших в процессе эволюции организма может выявить гены, необходимые для построения частей тела, гены, которые могут быть вовлечены в генетические нарушения человека. Например, мексиканская тетра - это пещерная рыба-альбинос, потерявшая зрение в процессе эволюции. Совместноеразмножение разных популяций этой слепой рыбы дало потомство с функциональными глазами, как разные мутации произошли в результате популяций, которые развивались в разных пещерах. Этопомогло идентифицировать гены, необходимые для зрения и пигментации.
Эволюционная теория имеет множество приложений в медицине. Многие человеческие болезни не являются статичными явлениями, но могут эволюционировать. Вирусы, бактерии, грибки и раковые образования развиваются и становятсяустойчивыми к иммунной защите хозяина, а также к фармацевтическим препаратам. Те же проблемы возникают в сельском хозяйстве с устойчивостью к пестицидам и гербицидам. Требуется более глубокое знание сложных сил, движущих эволюцию на молекулярном уровне, что мы стоим перед концом эффективных служб доступных антибиотиков, и для прогнозирования эволюции и эволюции наших патогенов и разработки стратегий для замедления или обхода.
В информатике моделирование эволюции с использованием эволюционных алгоритмов и искусственной жизни началось в 1960-х годах и было расширено за счет моделирования искусственногоотбора. Искусственная эволюция стала широко признанным методом оптимизации в результате работы Инго Рехенберга в 1960-х годах. Он использовал стратегии эволюции для решения сложных инженерных задач. Генетические алгоритмы, в частности, стали популярными благодаря написанию Джона Генри Холланда. Практические приложения также включают автоматическую эволюцию компьютерных программ. Эволюционные алгоритмы теперь используются для решениямногомерных задач более эффективно, чем программное обеспечение, созданное людьми-разработчиками, а также для оптимизации проектирования систем.
 Иисус эволюция получила широкое признание в 1870-х годах, карикатуры на Чарльза Дарвина с телом обезьяны илиобезьяны символизировали эволюцию.
Иисус эволюция получила широкое признание в 1870-х годах, карикатуры на Чарльза Дарвина с телом обезьяны илиобезьяны символизировали эволюцию. В 19 веке, особенно после публикации «Происхождения видов в мире» в 1859 году идея о том, что жизнь эволюционировала, былаактивным источником академических дебатов, предоставленных на философских, социальных и религиозных последствийх эволюции. Сегодня современный эволюционный синтез принимается подавляющим большинством ученых. Однако эволюция остается спорным понятием для некоторого теистов.
В то время как религии и конфессии помирились свои убеждения с эволюцией через такие понятия, как теистической эволюции, есть креационисты, считают, что эволюция противоречит мифамо сотворении, найденным в их религиях, и которые выдвигают различные возражения против эволюции. Как показали ответы на публикацию Остатков естественной истории сотворения мира в 1844 году, наиболее спорным аспектом эволюционной биологии является вывод >из эволюции человека о том, что люди имеют общее происхождение. с обезьянами и что умственные и моральные человеческие способности имеют те же естественные причины, которые и другиеунаследованные черты у животных. В некоторых странах, особенно в других странах, религиозный конфликт, сосредоточенный на политике и государственном образовании, эти противоречия между наукой и религией подпитывают нынешние противоречия между творением и эволюцией. В то время как другие научныеобласти, такие как космология и наука о Земле, также противоречат буквальным интерпретацией многих религиозных текстов, эволюционная биология встречаетбольшее сопротивление со стороны религиозных буквалистов.
Преподавание эволюции в американской классах биологии средней школы было необычно на протяжении большей части первой половины 20 века. Решение по делу Scopes Trial от 1925 года сделало этот предмет очень редким в американской биологиицелого поколения, но позже он был постепенно возвращен и стал юридически защищенным в 1968 году Эпперсон против Арканзаса решение. С тех пор конкурирующая религиозная вера вкреационизм была юридически запрещена в программах средней школы в различных решениях 1970-х и 1980-х годов, но вернулась в псевдонаучной форме как разумный замысел (ID), чтобы снова исключить его из дела 2005 Китцмиллер против школьного округа Дувр. Споры по поводу идейДарвина не вызвали значительных споров в Китае.