Показатель в эпидемиологии
Значения  общеизвестных инфекционных заболеваний (анализируется до любого социального вмешательства)
общеизвестных инфекционных заболеваний (анализируется до любого социального вмешательства)| Болезнь | Передача | |
|---|
| Корь | Аэрозоль | 12 –18 |
| Ветряная оспа (ветряная оспа) | Аэрозоль | 10–12 |
| Свинка | Респираторные капли | 10–12 |
| Полиомиелит | Фекально-оральный путь | 5–7 |
| Краснуха | Респираторные капли | 5–7 |
| Коклюш | Респираторные капли | 5,5 |
| Оспа | Респираторные капли | 3,5–6 |
| ВИЧ / СПИД | Биологические жидкости | 2–5 |
| SARS | Респираторные капли | 0,19–1,08 |
| COVID-19 | Респираторные капли. (Аэрозоль передача. исследуется) | 2–6 |
| Простуда | Респираторные капли | 2–3 |
| Дифтерия | Слюна | 1,7–4,3 |
| Грипп. (пандемический штамм 1918 г. ) | Респираторные капли | 1,4–2,8 |
| Эбола. (Вспышка лихорадки Эбола в 2014 г. ) | Биологические жидкости | 1,5–1,9 |
| Грипп. (пандемический штамм 2009 г. ) | Респираторные капли | 1,4–1,6 |
| Грипп. (сезонные штаммы) | Респираторные капли | 0,9–2,1 |
| MERS | Респираторные капли | 0,3–0,8 |
 Воспроизвести медиа
Воспроизвести медиа Видео, в котором обсуждается базовое количество воспроизведений (примерно через 4 минуты) и
коэффициент летальности в контексте
пандемии COVID-19.
в эпидемиология, базовое репродуктивное число или базовое репродуктивное число (иногда называемое базовым коэффициентом воспроизводства или базовым репродуктивным коэффициентом ), обозначенный (произносится как R ноль или R ноль), заражения можно рассматривать как ожидаемое число случаев, непосредственно вызванных одним случаем в популяции, где все люди восприимчивы к инфекции. Определение описывает состояние, в котором другие люди не инфицированы или иммунизированы (естественным путем или посредством вакцинации ). Некоторые определения, такие как определение Министерства здравоохранения Австралии, добавляют отсутствие «какого-либо преднамеренного вмешательства в передачу болезни». Базовый номер воспроизведения не следует путать с эффективным номером воспроизведения  (обычно пишется
(обычно пишется  [t для времени], иногда
[t для времени], иногда  ), который представляет собой количество наблюдений, сгенерированных в текущем состоянии генеральной совокупности, которое не обязательно должно быть незараженным. Также важно отметить, что - это безразмерное число, а не скорость, которая может иметь единицы времени или единицы времени, например время удвоения.
), который представляет собой количество наблюдений, сгенерированных в текущем состоянии генеральной совокупности, которое не обязательно должно быть незараженным. Также важно отметить, что - это безразмерное число, а не скорость, которая может иметь единицы времени или единицы времени, например время удвоения.
не является биологической константой для патогена, поскольку на него также влияют другие факторы, такие как условия окружающей среды и поведение инфицированного населения. Кроме того, значения обычно оцениваются на основе математических моделей, а оценочные значения зависят от используемой модели и значений других параметров. Таким образом, значения, приведенные в литературе, имеют смысл только в данном контексте, и рекомендуется не использовать устаревшие значения или сравнивать значения, основанные на разных моделях. действительно сами по себе не дают оценки того, насколько быстро инфекция распространяется среди населения.
Наиболее важные применения - это определение того, может ли возникающее инфекционное заболевание распространяться среди населения, и определение какая часть населения должна быть иммунизирована посредством вакцинации для искоренения болезни. В обычно используемых моделях заражения, когда  инфекция сможет начать распространяться среди населения, но только если
инфекция сможет начать распространяться среди населения, но только если  . Как правило, чем больше значение , тем сложнее контролировать эпидемию. Для простых моделей доля популяция, которую необходимо эффективно иммунизировать (то есть не восприимчивая к инфекции) для предотвращения устойчивого распространения инфекции, должна быть больше
. Как правило, чем больше значение , тем сложнее контролировать эпидемию. Для простых моделей доля популяция, которую необходимо эффективно иммунизировать (то есть не восприимчивая к инфекции) для предотвращения устойчивого распространения инфекции, должна быть больше  . И наоборот, доля населения, которая остается восприимчивой к инфекции в эндемическом равновесии, составляет
. И наоборот, доля населения, которая остается восприимчивой к инфекции в эндемическом равновесии, составляет  .
.
Базовое воспроизводство количество зависит от нескольких факторов, включая продолжительность заразности пораженных людей, t инфекционность микроорганизма и количество восприимчивых людей в популяции, с которыми контактируют инфицированные.
Содержание
- 1 История
- 2 Определения в конкретных случаях
- 2.1 Частота контактов и инфекционный период
- 2.2 С различными латентными периодами
- 2.3 Гетерогенные популяции
- 3 Методы оценки
- 3.1 Простая модель
- 3.2 Скрытый инфекционный период, изоляция после постановки диагноза
- 4 Эффективное число репродукции
- 5 Ограничения
- 6 В массовой культуре
- 7 См. Также
- 8 Примечания
- 9 Ссылки
- 10 Дополнительная литература
История
Корни базовой концепции воспроизводства можно проследить через работы Рональда Росс, Альфред Лотка и другие, но его первое современное применение в эпидемиологии было сделано Джорджем Макдональдом в 1952 году, который построил популяционные модели распространения малярии. В своей работе он назвал количественную базовую скорость воспроизведения и обозначил ее  . Обозначение величины ставкой может ввести в заблуждение, поскольку в таком случае «ставка» может быть неверно интерпретирована как число в единицу времени. «Число» или «соотношение» теперь предпочтительнее.
. Обозначение величины ставкой может ввести в заблуждение, поскольку в таком случае «ставка» может быть неверно интерпретирована как число в единицу времени. «Число» или «соотношение» теперь предпочтительнее.
Определения для конкретных случаев
Частота контактов и период заражения
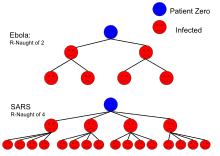
- среднее количество людей, инфицированных друг от друга человек. Например, у вируса Эбола
из двух, поэтому в среднем человек, болеющий Эболой, передаст его двум другим людям.
Предположим, что инфекционные люди совершают в среднем  вызывающих инфекцию контактов в единицу времени со средним инфекционным периодом
вызывающих инфекцию контактов в единицу времени со средним инфекционным периодом  . Тогда базовое число воспроизведения:
. Тогда базовое число воспроизведения:

Эта простая формула предлагает различные способы уменьшения и, в конечном итоге, распространение инфекции. Можно уменьшить количество вызывающих инфекцию контактов в единицу времени , уменьшив количество контактов в единицу времени (например, оставаясь дома, если инфекция требует контакт с другими для распространения) или доля контактов, вызывающих инфекцию (например, ношение какого-либо защитного снаряжения). Также возможно уменьшить инфекционный период путем скорейшего обнаружения, а затем изолирования, лечения или устранения (как часто бывает с животными) инфекционных особей.
С различными латентными периодами
Латентный период - это время перехода между событием заражения и проявлением болезни. В случае заболеваний с различными латентными периодами базовое число воспроизводств можно рассчитать как сумму чисел воспроизводства для каждого времени перехода в болезнь. Примером этого является туберкулез (ТБ). Воздуходувка и соавторы вычислили на основе простой модели TB следующий номер воспроизведения:

В их модели предполагается, что у инфицированных людей может развиться активный туберкулез либо путем прямого прогрессирования (болезнь развивается сразу после заражения), рассмотренного выше как БЫСТРЫЙ туберкулез, либо путем эндогенной реактивации. (заболевание развивается спустя годы после заражения), рассмотренное выше как МЕДЛЕННЫЙ туберкулез.
Гетерогенные популяции
В популяциях, которые не являются однородными, определение более тонкий. Определение должно учитывать тот факт, что типичный инфицированный человек может не быть обычным человеком. В качестве крайнего примера рассмотрим популяцию, в которой небольшая часть особей полностью смешивается друг с другом, а остальные особи все изолированы. Болезнь может распространяться в полностью смешанной части, даже если случайным образом выбранный человек приведет к менее чем одному вторичному случаю. Это связано с тем, что типичный инфицированный человек находится в полностью смешанной части и, таким образом, может успешно вызывать инфекции. В общем, если люди, инфицированные на ранней стадии эпидемии, в среднем либо более, либо менее вероятно передают инфекцию, чем люди, инфицированные на поздних этапах эпидемии, то вычисление должен учитывать это различие. Подходящим определением для в данном случае является «ожидаемое количество вторичных случаев, вызванных типичным инфицированным человеком в начале эпидемии».
Базовое число репродуктивных может быть вычислено как отношение известных коэффициентов с течением времени: если инфекционный человек контактирует с другими людьми в единицу времени, если предполагается, что все эти люди заразились болезнью, и если болезнь имеет средний инфекционный период 1 / γ, то базовое число воспроизведения просто R 0 = β / γ. Некоторые заболевания имеют несколько возможных латентных периодов, и в этом случае число воспроизводств для болезни в целом является суммой числа воспроизводств для каждого времени перехода в болезнь. Например, Blower et al. смоделируйте две формы туберкулезной инфекции: в быстром случае симптомы проявляются сразу после заражения; в медленном случае симптомы развиваются через годы после первоначального воздействия (эндогенная реактивация). Общее число воспроизведений является суммой двух форм сжатия: R 0 = R 0 + R 0.
Методы оценки
Базовое число воспроизведений может быть оценивается путем изучения подробных цепочек передачи или посредством геномного секвенирования. Однако чаще всего он рассчитывается с использованием эпидемиологических моделей. Во время эпидемии обычно известно количество диагностированных инфекций  за время
за время  . На ранних стадиях эпидемии рост является экспоненциальным, с логарифмической скоростью роста
. На ранних стадиях эпидемии рост является экспоненциальным, с логарифмической скоростью роста

Для экспоненциального роста  можно интерпретировать как совокупное количество диагнозов (включая выздоровевших) или текущее количество случаев инфекции; логарифмическая скорость роста одинакова для обоих определений. Чтобы оценить , необходимы предположения о временном промежутке между заражением и диагностикой, а также времени между заражением и началом распространения инфекции.
можно интерпретировать как совокупное количество диагнозов (включая выздоровевших) или текущее количество случаев инфекции; логарифмическая скорость роста одинакова для обоих определений. Чтобы оценить , необходимы предположения о временном промежутке между заражением и диагностикой, а также времени между заражением и началом распространения инфекции.
При экспоненциальном росте  связано со временем удвоения
связано со временем удвоения  как
как

Простая модель
Если человек после заражения заражает ровно новые особи только по прошествии ровно времени (последовательный интервал), затем количество инфекционных со временем растет как

или

Базовое дифференциальное уравнение сопоставления

или

В этом случае  или
или  .
.
Например, с  и
и  , мы найдем
, мы найдем  .
.
Если зависит от времени

показывает, что может быть важно сохранить  ниже 0, усредненное по времени, чтобы избежать экспоненциального роста.
ниже 0, усредненное по времени, чтобы избежать экспоненциального роста.
Скрытый инфекционный период, изоляция после постановки диагноза
В этой модели индивидуальная инфекция имеет следующие стадии:
- Открытая: человек инфицирован, но не имеет симптомов и еще не заражен другие. Средняя продолжительность открытого состояния
 .
. - Скрытая инфекция: человек инфицирован, не имеет симптомов, но заражает других. Средняя продолжительность латентного инфекционного состояния составляет
 . В течение этого периода человек заражает других лиц.
. В течение этого периода человек заражает других лиц. - изоляция после постановки диагноза: принимаются меры для предотвращения дальнейших инфекций, например путем изоляции инфицированный человек.
Это модель SEIR, и может быть записан в следующей форме

Это Метод оценки был применен к COVID-19 и SARS. Это следует из дифференциального уравнения для количества зараженных лиц  и количества латентных инфекционных лиц
и количества латентных инфекционных лиц  ,
,

Наибольшее собственное значение матрицы - это логарифмическая скорость роста , которая может решается для .
В частном случае  , это модель приводит к
, это модель приводит к  , что отличается от простой модели выше (
, что отличается от простой модели выше ( ). Например, с одинаковыми значениями и , мы найдем
). Например, с одинаковыми значениями и , мы найдем  , а не истинное значение
, а не истинное значение  . Разница обусловлена незначительной разницей в базовой модели роста; матричное уравнение выше предполагает, что недавно инфицированные пациенты уже вносят свой вклад в инфекции, тогда как на самом деле инфекции возникают только из-за числа инфицированных назад. Более правильное лечение потребует использования дифференциальных уравнений задержки.
. Разница обусловлена незначительной разницей в базовой модели роста; матричное уравнение выше предполагает, что недавно инфицированные пациенты уже вносят свой вклад в инфекции, тогда как на самом деле инфекции возникают только из-за числа инфицированных назад. Более правильное лечение потребует использования дифференциальных уравнений задержки.
Эффективное число воспроизводства
В действительности различные доли населения невосприимчивы к любому данному заболеванию в любой момент времени. Чтобы учесть это, используется эффективное число воспроизведения , обычно записываемое как , или среднее количество новых инфекций, вызванных одним инфицированным человеком в момент времени t в частично восприимчивой популяции. Его можно найти, умножив на долю S восприимчивой популяции. Когда доля иммунной популяции увеличивается (т. Е. Восприимчивая популяция S уменьшается) настолько, что падает ниже 1, «коллективный иммунитет "достигнуто, и количество случаев в популяции будет постепенно уменьшаться до нуля.
Ограничения
Использование в популярной прессе привело к недопониманию и искажению его значения. можно вычислить с помощью множества различных математических моделей. Каждый из них может дать различную оценку , которую необходимо интерпретировать в контексте этой модели. Следовательно, заразность различных инфекционных агентов нельзя сравнивать без пересчета с инвариантными предположениями. Значения для прошлых вспышек могут быть недопустимыми для текущих вспышек того же заболевания. Вообще говоря, может использоваться в качестве порога, даже если рассчитывается другими методами: если , вспышка исчезнет, и если  , вспышка будет расширяться. В некоторых случаях для некоторых моделей значения может по-прежнему приводить к самовоспроизводящимся вспышкам. Это особенно проблематично, если между хозяевами существуют промежуточные переносчики, такие как малярия. Поэтому сравнение значений из «Значения общеизвестных инфекционных заболеваний »следует вести с осторожностью.
, вспышка будет расширяться. В некоторых случаях для некоторых моделей значения может по-прежнему приводить к самовоспроизводящимся вспышкам. Это особенно проблематично, если между хозяевами существуют промежуточные переносчики, такие как малярия. Поэтому сравнение значений из «Значения общеизвестных инфекционных заболеваний »следует вести с осторожностью.
Хотя не может быть изменен путем вакцинации или других изменений в восприимчивости населения, его можно изменить путем физического дистанцирования и других мер воздействия. blic политика или социальное вмешательство. В совокупности большинство из них считаются нефармакологическими вмешательствами. Это создает некоторую путаницу, потому что не является константой; тогда как большинство математических параметров с индексами "ноль" являются константами.
зависит от многих факторов, многие из которых необходимо оценить. Каждый из этих факторов увеличивает неопределенность оценок . Многие из этих факторов не важны для информирования государственной политики. Следовательно, для государственной политики могут быть лучше использованы метрики, подобные , но которые проще оценить, например время удвоения или . период полураспада (t 1⁄2).
Методы, используемые для вычисления , включают функцию выживания, перестановка наибольшего собственного значения матрицы Якоби , метод следующего поколения, расчеты на основе внутренней скорости роста, существования эндемического равновесия, количества восприимчивых в эндемическом равновесии, средний возраст заражения и окончательное уравнение размера. Некоторые из этих методов согласуются друг с другом, даже если исходить из одной и той же системы дифференциальных уравнений. Еще меньшее число фактически вычисляет среднее количество вторичных инфекций. Поскольку редко наблюдается в полевых условиях и обычно рассчитывается с помощью математической модели, что сильно ограничивает его полезность.
В популярная культура
В фильме 2011 года Заражение, вымышленном триллере о медицинских катастрофах, расчеты блоггера для представлены, чтобы отразить прогрессирование смертельной вирусной инфекции от тематических исследований до пандемии. Изображенные методы были ошибочными.
См. Также
 Медицинский портал
Медицинский портал  Портал о коронавирусной болезни 2019
Портал о коронавирусной болезни 2019
Примечания
- Компартментные модели в эпидемиологии описывают динамику заболевания во времени в популяции восприимчивых (S), инфекционных (I) и выздоровевших ( R) люди, использующие модель SIR. Обратите внимание, что в модели SIR
 и разные количества - первое описывает количество выздоровевших при t = 0, тогда как второе описывает соотношение между частотой контактов и частотой выздоровления.
и разные количества - первое описывает количество выздоровевших при t = 0, тогда как второе описывает соотношение между частотой контактов и частотой выздоровления. - Согласно данным Провинциального центра по контролю и профилактике заболеваний Гуандун, «эффективный репродуктивное число (R или R e) чаще используется для описания передаваемости, которая определяется как среднее количество вторичных случаев, вызванных одним [sic] инфекционным случаем ». Например, по предварительной оценке во время продолжающейся пандемии эффективное репродуктивное число для SARS-CoV-2 было установлено равным 2,9, а для SARS - 1,77.
Ссылки
Дополнительная литература
- Heesterbeek, JAP (2002). «Краткая история и рецепт его вычисления». Acta Biotheoretica. 50 (3): 189–204. DOI : 10.1023 / A: 1016599411804. PMID 12211331. S2CID 10178944.
- Heffernan, J.M.; Smith, R.J.; Wahl, L.M. (октябрь 2005 г.). «Перспективы основного воспроизводственного соотношения». Журнал Интерфейса Королевского общества. 2 (4): 281–293. DOI : 10.1098 / RSIF.2005.0042. PMC 1578275. PMID 16849186.
- Джонс, Джеймс Холланд (1 мая 2007 г.). "Примечания к " (PDF). Получено 6 ноября 2018 г.
- Van Den Driessche, P.; Watmough, James (2008). «Дальнейшие примечания к основному репродуктивному числу». Математическая эпидемиология. Лекционные заметки по математике. 1945 . Pp. 159–178. doi : 10.1007 / 978-3-540-78911-6_6. ISBN 978-3-540-78910-9.

 Воспроизвести медиа Видео, в котором обсуждается базовое количество воспроизведений (примерно через 4 минуты) и коэффициент летальности в контексте пандемии COVID-19.
Воспроизвести медиа Видео, в котором обсуждается базовое количество воспроизведений (примерно через 4 минуты) и коэффициент летальности в контексте пандемии COVID-19.