
Войти
В филогенетике, базальный - это направление основания (или корня) корневого филогенетического дерева или кладограммы. Термин может более строго применяться только к узлам, смежным с корнем, или более свободно к узлам, рассматриваемым как близкие к корню. Каждый узел в дереве соответствует кладе ; то есть кладу C можно описать как базальную в пределах более крупной клады D, если ее корень напрямую связан с корнем D. Термины глубокое ветвление или раннее ветвление аналогичны по значению.
Хотя всегда должно быть две или более одинаковых базальных клады, прорастающих из корня каждой кладограммы, эти клады могут сильно различаться по таксономическому рангу, видовому разнообразию или и то и другое. Если C является базальной кладой внутри D, которая имеет самый низкий ранг среди всех базальных клад внутри D, C может быть описан как базальный таксон этого ранга внутри D. Концепция 'ключевой инновации 'подразумевает некоторую степень корреляции между эволюционными инновациями и диверсификацией. Однако такая корреляция не делает данный случай предсказуемым, поэтому наследственные признаки не следует вменять членам базальной клады с меньшим видовым разнообразием без дополнительных доказательств.
В целом, клады A более базальна, чем клады B, если B является подгруппой сестринской группы A или самой A. Внутри больших групп термин «базальный» может использоваться в широком смысле для обозначения «ближе к корню, чем подавляющее большинство», и в этом контексте может возникнуть такая терминология, как «очень базальный». «Основная клада» - это клада, представляющая все, кроме базальной клады (ки) самого низкого ранга в пределах более крупной клады; например, core eudicots.
Базальная группа в более строгом смысле образует сестринскую группу для остальной части большей клады, как в следующем случае:
| Базальная клада # 2 |
Хотя базальную кладу в такая кладограмма, уместность такой идентификации зависит от точности и полноты схемы. В этом примере предполагается, что конечные ветви кладограммы изображают все существующие таксоны данного ранга внутри клады; в противном случае диаграмма может оказаться весьма обманчивой. Кроме того, эта квалификация не гарантирует, что представлено разнообразие вымерших таксонов (которые могут быть плохо изучены).
В филогенетики, термин базально может быть объективно применены к кладам организмов, но, как правило, применяются избирательно и более спорна к группам или родословных как полагает, обладают родовыми символами, или к таким предположительно наследственные черты сами по себе. При описании символов «предковый» или «плезиоморфный » предпочтительнее, чем «базальный» или «примитивный », последний из которых может иметь ложные коннотации неполноценности или несложности.
Несмотря на повсеместное использование базального, некоторые систематики считают, что его применение к существующим группам не нужно и вводит в заблуждение. Этот термин чаще применяется, когда одна ветвь (та, которая считается «базальной») менее разнообразна, чем другая ветвь (это ситуация, в которой можно было бы ожидать найти базальный таксон более низкого минимального ранга). Этот термин может быть двусмысленным, поскольку он также относится к направлению корня дерева, которое представляет гипотетического предка; это, следовательно, может неточно означать, что сестринская группа более богатой видами клады демонстрирует наследственные черты. Существующая базальная группа может напоминать, а может и не напоминать последнего общего предка более крупной клады в большей степени, чем другие группы, и отделена от этого предка на такое же количество времени, что и все другие существующие группы. Однако есть случаи, когда необычно маленький размер сестринской группы действительно коррелирует с необычным количеством наследственных черт, как в Amborella (см. Ниже). Другими известными примерами этого явления являются яйцеклад и лактация без сосков монотрем, базальная клада млекопитающих, насчитывающая всего пять видов, и архаическая анатомия tuatara, базальная клады лепидозавров с одним видом.
 Amborella trichopoda, самый базальный из существующих покрытосеменных
Amborella trichopoda, самый базальный из существующих покрытосеменных цветковых растений семейства Amborellaceae, ограниченная Новой Каледонией в юго-западной части Тихого океана, представляет собой базальную кладу современных покрытосеменных, состоящую из большинства основных видов, родов, семейств и отрядов внутри группы (из общего числа около 250 000 видов покрытосеменных). Признаки Amborella trichopoda считаются важными для понимания эволюции цветковых растений; например, у него есть «самая примитивная древесина (состоящая только из трахеид ) из всех живых покрытосеменных», а также «простые отдельные части цветка с неопределенным числом и незапечатанные плодолистики». Однако эти признаки представляют собой смесь архаичных и апоморфных (производных) признаков, которые были отсортированы только путем сравнения с другими покрытосеменными и их положениями в филогенетическом дереве (летопись окаменелостей могла потенциально также может быть полезным в этом отношении, но отсутствует в данном случае). Кладограмма ниже основана на Ramírez-Barahona et al. (2020), при этом подсчет видов взят из указанного источника.
| 'Базальные покрытосеменные ' |
Среди человекообразных обезьян гориллы (восточные и западные) являются родственной группой обыкновенным шимпанзе, бонобо и людям. Эти пять видов образуют кладу, подсемейство Homininae (африканские обезьяны), из которых гориллы являются основным родом. Однако, если анализ не ограничивается родами, клады Homo plus Pan также являются базальными.
 Филогенетическое дерево сумчатых, полученное на основе данных ретропозона, показывает базальное положение южноамериканских Didelphimorphia в пределах Marsupialia, а базальное положение южноамериканского Dromiciops внутри австралийской австралидельфии.
Филогенетическое дерево сумчатых, полученное на основе данных ретропозона, показывает базальное положение южноамериканских Didelphimorphia в пределах Marsupialia, а базальное положение южноамериканского Dromiciops внутри австралийской австралидельфии.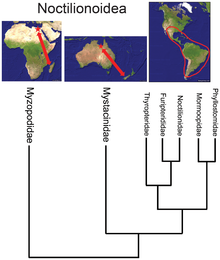 Взаимосвязь биогеографии и филогении надсемейства летучих мышей Noctilionoidea, выведенная из данных последовательностей ядерной ДНК, показывающая базальное положение малагасийских семейство Myzopodidae. Места, где есть только ископаемые, обозначены красными звездочками.
Взаимосвязь биогеографии и филогении надсемейства летучих мышей Noctilionoidea, выведенная из данных последовательностей ядерной ДНК, показывающая базальное положение малагасийских семейство Myzopodidae. Места, где есть только ископаемые, обозначены красными звездочками. | Homininae |
| ||||||||||||||||||||||||
Кроме того, орангутаны являются сестринской группой по отношению к Homininae и являются основным родом в семействе больших обезьян Hominidae в целом.
| Hominidae |
| ||||||||||||||||||
Подсемейства Homininae и Ponginae являются базальными внутри Hominidae, но, учитывая, что небазальных подсемейств на кладограмме нет, маловероятно, что этот термин будет применяться к обоим. В общем, утверждение о том, что одна группа (например, орангутанги) является основной или ответвляется первой внутри другой группы (например, Hominidae), может не иметь смысла, если только соответствующий таксономический уровень (уровни) (род, в данном случае) указан. Если этот уровень не может быть указан (т. Е. Если рассматриваемая клада не имеет ранга), может потребоваться более подробное описание соответствующих сестринских групп.
В этом примере орангутаны отличаются от других родов своим азиатским ареалом. Этот факт в сочетании с их базовым статусом указывает на то, что самый недавний общий предок современных человекообразных обезьян мог быть евразийцем (см. Ниже), что согласуется с другими доказательствами. (Конечно, меньшие обезьяны полностью азиатские.) Орангутаны также отличаются от африканских обезьян своим более древесным образом жизни - черта, обычно считающаяся у обезьян наследственной.
Учитывая, что самый глубокий филогенетический раскол в группе, вероятно, произошел в начале ее истории, идентификация наиболее базальных субкладов в широко рассредоточенном таксоне или кладе может дать ценные результаты. понимание региона его происхождения. В некоторых ситуациях, когда это может быть неочевидно, также можно сделать вывод о направлении миграции от района происхождения (как в случаях Amaurobioides и Noctilionoidea ниже). Примеры включают: