
Войти
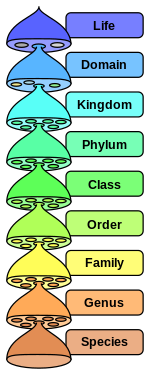 Иерархия восьми основных таксономических рангов по биологической классификации. Промежуточные второстепенные ранги не показаны.
Иерархия восьми основных таксономических рангов по биологической классификации. Промежуточные второстепенные ранги не показаны. Бактериальная таксономия - это таксономия, т. Е. ранг классификация бактерий.
в по научной классификации, установленной Карлом Линнеем, каждый вид должен быть отнесен к роду (бинарная номенклатура ), который, в свою очередь, является нижним уровнем иерархии ранги (семья, подотряд, порядок, подкласс, класс, подразделение / тип, королевство и домен). В принятой в настоящее время классификации жизни существует три области (эукариоты, бактерии и археи ), которые, с точки зрения таксономии, несмотря на следование одним и тем же принципам, имеют несколько различных соглашений между собой и между своими подразделениями, поскольку они изучаются разными дисциплинами (ботаника, зоология, микология и микробиология). Например, в зоологии есть типовые образцы, а в микробиологии - типовые.
Прокариоты имеют много общих черт, таких как отсутствие ядерной мембраны, одноклеточность, деление на бинарное деление и, как правило, небольшой размер. Различные виды различаются между собой по нескольким характеристикам, что позволяет их идентифицировать и классифицировать. Примеры включают:
Бактерии впервые были обнаружены Антони ван Левенгук в 1676 г., используя однообъективный микроскоп собственной конструкции п. Он назвал их «анималкулы » и опубликовал свои наблюдения в серии писем в Королевское общество.
. Ранние описанные роды бактерий включают Vibrio и Monas, автор О. Ф. Мюллер (1773, 1786), затем классифицированный как Infusoria (однако многие виды, ранее входившие в эти роды, сегодня считаются протистами); Polyangium, автор H. F. Link (1809), первая бактерия, известная до сих пор; Серратия, автор Бизио (1823 г.); и Spirillum, Spirochaeta и Bacterium, от Эренберга (1838).
Термин Bacterium, введенный как род Эренберга в 1838 году, стал универсальным для палочковидных клеток.
 Древо жизни в Generelle Morphologie der Organismen (1866)
Древо жизни в Generelle Morphologie der Organismen (1866) Бактерии сначала были классифицированы как растения, составляющие класс Schizomycetes, которые вместе с Schizophyceae (сине-зеленые водоросли / цианобактерии) сформировали тип Schizophyta.
Haeckel в 1866 году поместил группу в тип Moneres ( от μονήρης: простой) в королевстве Протиста и определяет их как полностью бесструктурные и однородные организмы, состоящие только из кусочка плазмы. Он разделил этот тип на две группы:
Классификация Фердинанда Кона (1872) оказала влияние на девятнадцатого века и выделил шесть родов: Micrococcus, Bacterium, Bacillus, Vibrio, Spirillum и Spirochaeta.
Группа была позже реклассифицирована как Prokaryotes Chatton.
Классификация Cyanobacteria (в просторечии «сине-зеленые водоросли») велась между водоросли или бактерии (например, Геккель отнес Nostoc к типу Archephyta of Algae).
в 1905 году Эрвин Ф. Смит принял 33 действительных различных названия бактериальных родов и более 150 неправильных названий, а Вюйлемин в исследовании 1913 года пришел к выводу, что все виды бактерий должны попадать в роды, Streptococcus, Klebsiella, Planomerista, Neisseria, Sarcina, Metabacterium, Clostridium, Serratia, Bacterium и Spirillum.
Кон распознал четыре трибы : Spherobacteria, Microbacteria, Desmobacteria и Spirobacteria. Станье и ван Нил признали королевство Монера с двумя типами: Myxophyta и Schizomycetae, последняя включает классы Eubacteriae (три отряда), Myxobacteriae (один отряд) и Spirochetae (один отряд). Биссет выделил 1 класс и 4 порядка: Eubacteriales, Actinomycetales, Streptomycetales и Flexibacteriales. Система Уолтера Мигулы, которая была наиболее широко принятой системой своего времени и включала все известные тогда виды, но была основана только на морфологии, включала три основные группы Coccaceae, Bacillaceae и Spirillaceae, а также Trichobacterinae для нитчатых бактерий. Орла-Йенсен установил два отряда: Cephalotrichinae (семь семейств) и Peritrichinae (предположительно, только с одним семейством). Bergey et al. представили классификацию, которая в целом соответствует Заключительному отчету Общества американских бактериологов за 1920 г. (Winslow et al.), в котором класс шизомицетов разделен на четыре отряда: Myxobacteriales, Thiobacteriales, Chlamydobacteriales и Eubacteriales, причем пятая группа состоит из четырех родов, считающихся промежуточными. между бактериями и простейшими: Spirocheta, Cristospira, Saprospira и Treponema.
Однако разные авторы часто переклассифицировали роды из-за отсутствия видимых признаков, что приводило к плохому состоянию, которое было резюмировано в 1915 году Робертом Эрлом Бьюкененом. К тому времени вся группа получила разные звания и названия разными авторами, а именно:
Кроме того, семейства, на которые был разделен класс, менялись от автора к автору, и для некоторых, таких как Zipf, имена были на немецком, а не на латыни.
Первое издание Бактериологического кодекса в 1947 году разрешило несколько проблем.
А. Система Р. Прево) имела четыре подтипы и восемь классов, а именно:
| Linnaeus. 1735 | Haeckel. 1866 | Chatton. 1925 | Коупленд. 1938 | Уиттакер. 1969 | Вез и др.. 1990 | Кавальер-Смит. 1998 | Кавалер-Смит. 2015 |
|---|---|---|---|---|---|---|---|
| 2 королевства | 3 королевства | 2 империи | 4 королевства | 5 королевств | 3 домена | 2 империи, 6 королевств | 2 империи, 7 королевств |
| (не лечится) | Протиста | Прокариота | Монера | Монера | Бактерии | Бактерии | Бактерии |
| Археи | Археи | ||||||
| Eukaryota | Protoctista | Protista | Eucarya | Protozoa | Protozoa | ||
| Chromista | Chromista | ||||||
| Vegetabilia | Plantae | План tae | Plantae | Plantae | Plantae | ||
| Fungi | Fungi | Fungi | |||||
| Animalia | Animalia | Animalia | Animalia | Animalia | Animalia |
Несмотря на отсутствие согласия по основным подгруппам бактерий, окрашивание по Граму чаще всего использовалось в качестве инструмента классификации. Следовательно, до появления молекулярной филогении Kingdom Prokaryotae была разделена на четыре подразделения. Схема классификации все еще формально сопровождалась руководством Берджи по систематической бактериологии для порядка
 Филогенетическое древо, показывающее взаимосвязь между археями и другими формами жизни. Эукариоты окрашены в красный цвет, археи - в зеленый, а бактерии - в синий. По материалам Ciccarelli et al.
Филогенетическое древо, показывающее взаимосвязь между археями и другими формами жизни. Эукариоты окрашены в красный цвет, археи - в зеленый, а бактерии - в синий. По материалам Ciccarelli et al. Вёзе утверждал, что бактерии, археи и эукариоты представляют собой отдельные линии происхождения, которые на раннем этапе разошлись с наследственной колонией организмов. Однако некоторые биологи утверждают, что археи и эукариоты произошли от группы бактерий. В любом случае считается, что вирусы и археи начали отношения примерно два миллиарда лет назад, и что совместная эволюция могла происходить между членами этих групп. Возможно, что последним общим предком бактерий и архей был термофил, что повышает вероятность того, что более низкие температуры являются «экстремальной средой» в терминах архей, а организмы, живущие в более прохладной среде, появились только позже. Поскольку археи и бактерии не более связаны друг с другом, чем с эукариотами, единственное сохранившееся значение термина «прокариот» - «не эукариот», что ограничивает его ценность.
С улучшенными методиками стало ясно, что метаногенез бактерии были глубоко разными и (ошибочно) считались реликтами древних бактерий, таким образом Карл Вёзе, считающийся предшественником революции молекулярной филогении, определил три основные линии происхождения: архебактерии, эубактерии и Уркариоты, последние теперь представлены нуклеоцитоплазматическим компонентом эукариот. Эти линии были формализованы в ранговый домен (regio на латыни), который разделил жизнь на 3 домена: эукариоты, археи и бактерии.
В 1987 году Карл Вёзе разделил эубактерии на 11 делений основаны на последовательностях 16S рибосомной РНК (SSU), которые с некоторыми добавлениями все еще используются сегодня.
Хотя трехдоменная система широко распространена, некоторые авторы выступали против этого по разным причинам.
Одним из видных ученых, выступающих против трехдоменной системы, является Томас Кавальер-Смит, который предположил, что археи и эукариоты (Neomura ) происходят от грамположительных бактерий. (Posibacteria), которые, в свою очередь, происходят от грамотрицательных бактерий (Negibacteria) на основании нескольких логических аргументов, которые весьма противоречивы и обычно не принимаются во внимание сообществом молекулярной биологии (см. Комментарии рецензентов, например, Эрик Баптест является «агностиком» в отношении выводов.) и часто не упоминаются в обзорах (например) из-за субъективного характера сделанных предположений.
Однако, несмотря на наличие множества статистически подтвержденных исследований, направленных на укоренение дерева жизни между Бактериями и Неомура с помощью различных методов, в том числе тех, которые невосприимчивы к ускоренной эволюции - что, по словам Кавалье-Смита, является источником предполагаемой ошибки в молекулярных методах - есть несколько исследований, которые показали d различные выводы, некоторые из которых помещают корень в тип Firmicutes с вложенными археями.
Молекулярная таксономия Радхи Гупты, основанная на консервативных сигнатурных последовательностях белков, включает монофилетическую грамотрицательную кладу, монофилетическую грамположительную кладу и полифилетическая археота, полученная из грамположительных. Молекулярный анализ Хори и Осавы показал связь между метабактериями (= Archeota) и эукариотами. Единственный кладистический анализ бактерий, основанный на классических доказательствах, в значительной степени подтверждает результаты Гупты (см. всеобъемлющую мегатаксономию ).
Джеймс Лейк представил 2 основных устройства царства (Parkaryotae + эукариоты и эоциты + Karyotae) и предложил схему 5 основных царств (Eukaryota, Eocyta, Methanobacteria, Halobacteria и Eubacteria) на основе рибосомальной структуры и 4 основных схема царства (Eukaryota, Eocyta, Methanobacteria и), бактерии классифицируются по трем основным биохимическим нововведениям: фотосинтез (Photocyta), метаногенез (Methanobacteria), и (Eocyta ). Он также обнаружил доказательства того, что грамотрицательные бактерии возникли в результате симбиоза между двумя грамположительными бактериями.
Классификация - это группирование организмов в все более инклюзивные группы на основе филогении и фенотипа., в то время как номенклатура - это применение формальных правил для наименования организмов.
Несмотря на отсутствие официальной и полной классификации прокариот, названия (номенклатура), даваемые прокариотам, регулируются Международный кодекс номенклатуры бактерий (Бактериологический кодекс ), книга, которая содержит общие соображения, принципы, правила и различные примечания, а также дает советы, аналогичные кодам номенклатуры других группы.
Таксоны, которые были правильно описаны, рассмотрены в руководстве Бергей по систематической бактериологии, которое направлено на помощь в идентификации видов и считается высшим авторитетным источником. Доступна онлайн-версия таксономического обзора бактерий и архей (TOBA) [1].
Список названий прокариот, стоящих в номенклатуре (LPSN) - это онлайн-база данных, которая в настоящее время содержит более двух тысяч принятых названий с их ссылки, этимологии и различные примечания.
The International Journal of Systematic Bacteriology / Международный журнал систематической и эволюционной микробиологии (IJSB / IJSEM) - это рецензируемый журнал, который действует как официальный международный форум по публикации новых таксонов прокариот. Если вид опубликован в другом рецензируемом журнале, автор может подать запрос в IJSEM с соответствующим описанием, которое, если оно верное, приведет к тому, что новый вид будет включен в Список валидации IJSEM.
Коллекции микробных культур представляют собой хранилища штаммов, которые направлены на их защиту и распространение. Основными из них являются:
| Коллекция Акроним | Название | Местоположение |
|---|---|---|
| ATCC | Американская коллекция типовых культур | Манассас, Вирджиния |
| NCTC | Национальная коллекция типовых культур | Общественное здравоохранение Англии, Соединенное Королевство |
| BCCM | Скоординированная коллекция микроорганизмов Бельгии | Гент, Бельгия |
| CIP | Collection d'Institut Pasteur | Париж, Франция |
| DSMZ | Deutsche Sammlung von Mikroorganismen und Zellkulturen | Брауншвейг, Германия |
| JCM | Коллекция микроорганизмов Японии | Сайтама, Япония |
| NCCB | Коллекция культур Нидерландов бактерий | Утрехт, Нидерланды |
| NCIMB | Национальная коллекция промышленных, морских и пищевых бактерий | Абердин, Шотландия |
| ICMP | Международная коллекция микроорганизмов из растений | Окленд, Новая Зеландия |
| CECT | Валенсия, Испания |
Бактерии сначала были классифицированы основываясь исключительно на их форме (вибрионы, палочки, кокки и т. д.), наличии эндоспор, окраске по Граму, аэробных условиях и подвижности. Эта система изменилась с изучением метаболических фенотипов, в которых использовались метаболические характеристики. В последнее время, с появлением молекулярной филогении, для идентификации видов используются несколько генов, наиболее важным из которых является ген 16S рРНК, за которым следует ген 23S, gyrB и др. для подтверждения лучшего разрешения. Самый быстрый способ определить соответствие изолированного штамма какому-либо виду или роду сегодня - это амплификация его гена 16S с помощью универсальных праймеров, секвенирование ампликона размером 1,4 КБ и отправка его в специализированную базу данных идентификации в Интернете, а именно [ 2], которые выравнивают последовательность с другими последовательностями 16S, используя infernal, вторичная структура, основанная на глобальном выравнивании, или, которая выравнивает последовательности через SINA (инкрементный выравниватель SILVA), который выполняет локальное выравнивание затравки и расширяет его [3].
Существует несколько методов идентификации:
Минимальные стандарты для привлечение нового вида зависят от того, к какой группе принадлежит этот вид. Cf
Candidatus - это компонент таксономического названия бактерии, которая не может находиться в коллекции бактериологических культур. Например, «Candidatus Pelagibacter ubique "
Бактерии делятся бесполым путем и по большей части не проявляют регионализма («Все везде »), поэтому концепция видов, который лучше всего подходит для животных, становится исключительно предметом суждения.
Число названных видов бактерий и архей (примерно 13 000) на удивление мало, их раннюю эволюцию, генетическое разнообразие и место обитания во всех экосистемах. Горизонтальный перенос генов, размывающий различие видов, предварительное условие для использования в культуре (предварительное условие для новых видов, см. Выше) и обширный горизонтальный перенос генов, размывающий различные виды.
Наиболее общепринятым определением является определение многофазного вида, учитывает как фенотипические, так и генетические различия. Однако более быстрый диагностический специальный порог для отдельных видов меньше 70% гибридизация ДНК-ДНК, что соответствует менее 97% идентичности ДНК 16S. Было отмечено, что если бы это было применено к классификации животных, приматы отряда были бы одним видом. По этой причине были предложены более строгие определения видов, основанные на полногеномных последовательностях.
В идеале таксономическая классификация должна отражать эволюционную историю таксонов, то есть филогению. Хотя существуют некоторые исключения, когда фенотип различается в группе, особенно с медицинской точки зрения. Ниже приведены некоторые примеры проблемных классификаций.
В семействе Enterobacteriaceae класса Gammaproteobacteria, виды из рода Shigella (, S. Flexneri, S. boydii, S. sonnei ) с эволюционной точки зрения являются штаммами вида Escherichia coli (полифилетический), но из -за генетических различий вызывает различные медицинские состояния в случае патогенных штаммов. Escherichia coli имеют плохо классифицируется как вид, поскольку некоторые штаммы только 20% общего генома. Он настолько задействован, ему следует дать более высокий таксономический рейтинг. Однако из-за медицинских условий, связанных с этим видом, он не будет изменен, чтобы избежать путаницы в медицинском контексте.
Подобным образом виды Bacillus (= тип Firmicutes ) принадлежат к «группе B. cereus» (B. anthracis, B. cereus, B. thuringiensis, B. mycoides, B. pseudomycoides, B. weihenstephanensis и) имеют 99-100% схожую последовательность 16S рРНК (97% - это обычно используемый адекватный видовой предел) и являются полифилетическими, но по медицинским причинам (сибирская язва и т. Д.) Остаются отдельными.
Yersinia pestis в действительности является штаммом Yersinia pseudotuberculosis, но с островком патогенности, которая вызывает совершенно иную патологию (черная чума и туберкулезный симптомы соответственно), которые возникли от 15000 до 20000 лет назад.
В отряде гаммапротеобактерий Pseudomonadales, род Azotobacter и на самом деле являются членами родов Pseudomonas, но были неправильно классифицированы из-за способности фиксировать азот и большой тип размера Pseudomonas, что затрудняет классификацию. Вероятно, это будет исправлено в ближайшем будущем.
Другим примером большого рода вложенных родами является род Bacillus, в котором роды Paenibacillus и Brevibacillus являются вложенными кладами. В настоящее время геномных данных для полного и эффективного исправления таксономических ошибок Bacillus.
На основании молекулярных данных было показано, что род Agrobacterium гнездится в Rhizobium, а виды Agrobacterium переведены в род Rhizobium (в результате получено следующее сравнение.: Rhizobium radiobacter (ранее известная как A. tumefaciens), R. rhizogenes, R. rubi, R. undicola и R. vitis) Учитывая патогенную природу видов Agrobacterium для растений, было предложено сохранить род Agrobacterium, и последний был аргументирован контраргументом
Таксономические названия пишутся курсивом (или подчеркнуты, если написаны от руки) с большой первой буквой, за исключением эпитетов для видов и подвидов. Несмотря на то, что это распространено в зоологии, тавтонимы (например, Bison bison) неприемлемы, а названия таксонов, используются в зоологии, ботанике или микологии, не могут быть повторно использованы для бактерий (ботаника и зоология имеют общие названия).
Номенклатура - это набор правил и соглашений, регулирующих названия таксонов. Разница в номенклатуре между различными царствами / доменами в.
Для бактерий допустимые имена должны иметь латинское или неолатинское имя и могут использовать только основные латинские буквы (w и jительно, см. История латинского алфавита для них), следовательно, дефисы, акценты и другие буквы не принимаются и должны быть правильно транслитерированы (например, ß = ss). Древнегреческий язык, написанный греческим алфавитом, должен быть транслитерирован в латинский алфавит.
Когда последнее не начинается с гласной, требуется соединительная гласная в зависимости от происхождения слова, независимо от того, что происходит за ним, если последнее не начинается с гласной. гласная. Если первое соединение латинское, то соединительная гласная - -i-, тогда как если первое соединение - греческое, соединяющая гласная - -o-.
Этимологию имен см. LPSN.
Для прокариот (бактерий и архей) ранговое царство не используется (хотя некоторые авторы называют филы царствами)
Если новый илиенный вид помещен в новые ранги, в соответствии с Правилом 9 Бактериологического кодекса название формируется путем добавления соответствующего суффикса к основанию названия типового рода. Для подкласса и класса обычно следует рекомендация от, что приводит к следующему множественному множеству, однако некоторые имена не следуют этому и учитывают греко-латинскую грамматику (например, женское множественное число Thermotogae, Aquificae и Chlamydiae, множественное число мужских Chloroflexi, Bacilli и Deinococci и греческое множественное число Spirochaetes, Gemmatimonadetes и Chrysiogentes ).
| Ранг | Суффикс | Пример |
|---|---|---|
| Род | -ae | (Elusimicrobiae) |
| Subtribe (вышедшее из употребления) | -inae | (Elusimicrobiinae) |
| Племя (вышедшее из употребления) | -eae | (Elusimicrobiieae) |
| Подсемейство | -oideae | (Elusimicrobioideae) |
| Семейство | -aceae | Elusimicrobiaceae |
| Подотряд | -ineae | (Elusimicrobineae) |
| Порядок | -ales | Elusimicrobiales |
| П одкласс | -idae | (Elusimicrobidae) |
| Класс | -ia | El usimicrobia |
| Тип | см. текст | Elusimicrobia |
Типы не подпадают под Бактериологический кодекс, однако научное сообщество обычно используется Таксономия Ncbi и Lpsn, в которой название типа, как правило, является многочисленным числом от типового рода, за исключением Фирмикуты, цианобактерии и протеобактерии, названия которых не имеют названия от рода. Высшие таксоны, предложенные Кавалье-Смитом, обычно игнорируются сообществом молекулярной филогении (например, см. Выше).
Для архей используется суффикс -археота. Для бактериального типа было предложено использовать суффикс -бактерии для типа.
Следовательно, для основного типа название типа такое же, как и у первого описанного класса:
, тогда как для других, где суффикс -ia для класса используется независимо от грамматики, они различаются:
Исключением Тип является типом Deinococcus - Thermus, который содержит родов через дефис - допустимые названия принимаются только латинскими буквами без ударения, но официально не признаются. логический код, чтобы указать -aeota как окончание бактериального типа и это имена производными от класса типа внутри филума. Для этого потребуются следующие изменения:
Некоторые виды названы в честь людей, либо открывателей, либо известных людей в области микробиологии, например Salmonella после DE Лосось, открывший его (хотя и «Bacillus typhi»).
Для общего эпитета все имена, производные от людей, должны быть в женском именительном падеже, либо путем изменения окончания на -a, либо на уменьшительное -ella, в зависимости от имени.
Для конкретного эпитета имена могут быть преобразованы в любую форму прилагательного (добавление -nus (m.), -na (f.), -num (n.) в зависимости от пола названия рода) или родительного падежа латинизированного названия.
Многие виды (специфический эпитет) названы по месту, где они присутствуют или обнаружены ( например). Их имена образованы путем образования прилагательного путем соединения названия местности с окончанием -ensis (m. Или f.) Или ense (n.) В соответствии с полом названия рода, если только для места не существует классического латинского прилагательного.. Однако названия мест не должны использоваться в качестве существительных в родительном падеже.
Несмотря на то, что некоторые гетеро / гомогенные колонии или биопленки бактерий имеют названия на английском языке (например, зубной налет или звездчатое желе ), ни один вид бактерий не имеет общеупотребительного / банального / общепринятого названия на английском языке.
Для имен в единственном числе нельзя использовать множественное число (singulare tantum ), так как это подразумевает наличие нескольких групп с одним и тем же ярлыком, а не нескольких членов этой группы (по аналогии, на английском языке, стулья и столы - это типы мебели, которые нельзя использовать во множественном числе «мебель» для описания обоих элементов), и наоборот, имена во множественном числе имеют форму множественного числа tantum. Однако частичное исключение из этого правила составляет использование имен на местном языке. Однако, чтобы избежать повторения таксономических названий, которые нарушают поток прозы, часто используются и рекомендуются народные названия представителей рода или более высоких таксонов, которые формируются путем написания названия таксона в латинском регистре предложения («стандарт» в MS Office), поэтому трактует имя собственное как английское нарицательное (например, сальмонеллы), хотя есть некоторые споры о грамматике множественного числа, которое может быть правильным множественным числом путем добавления - (e) s (сальмонеллы) или использования древнегреческой или латинской формы множественного числа (неправильное множественное число ) существительного (сальмонеллы); последнее проблематично, так как множественное число от будет -bacteres, а множественное число от myces (NL masc. n. от греч. masc. n. mukes) - это mycetes.
Существуют обычаи для некоторых имен, таких как поскольку те, которые заканчиваются на -monas, преобразуются в -монаду (одна псевдомонада, две аэромонады и не -монады).
Бактерии, которые являются этиологической причиной заболевания, часто обозначаются названием болезни, за которым следует описывающее существительное (бактерия, бацилла, кокк, возбудитель или название их типа), например бактерии холеры (Vibrio cholerae ) или спирохеты болезни Лайма (Borrelia burgdorferi ), обратите внимание также на риккетсиоз (Rickettsia akari ) (подробнее см.).
Трепонема преобразована в трепонему, и множественное число - это трепонемы, а не трепонемы.
Некоторые необычные бактерии имеют особые названия, такие как овал Куина (Quinella ovalis ) и квадрат Уолсби (Haloquadratum walsbyi ).
До появления молекулярной филогении многие более высокие таксономические группы имели только тривиальные названия, которые используются до сих пор, некоторые из которых являются полифилетическими, например Rhizobacteria. Некоторые более высокие таксономические тривиальные названия: