Войти
| Просмотр / редактирование Человека | Просмотр / редактирование мыши |
BRAF - человеческий ген, который кодирует белок , называемый B-Raf. Ген также упоминается как протоонкоген B-Raf и v-Raf гомолог вирусного онкогена B мышиной саркомы, в то время как белок более формально известен как серин / треонин-протеинкиназа B-Raf .
Белок B-Raf участвует в передаче сигналов внутри клеток, которые участвуют в управлении ростом клеток. В 2002 году было показано, что он ошибочен (мутировал ) в некоторых человеческих раковых опухолях.
. Некоторые другие наследственные мутации BRAF вызывают врожденные дефекты.
Разработаны препараты для лечения рака, вызванного мутациями BRAF. Два из этих препаратов, вемурафениб и дабрафениб, одобрены FDA для лечения меланомы на поздней стадии. Вемурафениб был первым одобренным лекарством, появившимся в результате открытия лекарств на основе фрагментов.
B-Raf является членом Raf-киназ семейства протеинкиназ передачи сигнала . Этот белок играет роль в регуляции MAP-киназы / ERKs сигнального пути, который влияет на деление клеток, дифференцировку и секреция.
B-Raf представляет собой 766- аминокислоту, регулируемую передачу сигнала серин / треонин- специфическая протеинкиназа. Вообще говоря, он состоит из трех консервативных доменов, характерных для семейства киназ Raf : консервативная область 1 (CR1), Ras - GTP -связывающий саморегулирующий домен, консервативная область 2 (CR2), шарнирная область, обогащенная серином, и консервативная область 3 (CR3), каталитический домен протеинкиназы, который фосфорилирует консенсусную последовательность на белковых субстратах. В своей активной конформации B-Raf образует димеры за счет водородных связей и электростатических взаимодействий своих киназных доменов.
Консервированная область 1 автоингибирует Киназный домен B-Raf (CR3), так что передача сигналов B-Raf регулируется, а не конститутивно. Остатки 155–227 составляют Ras -связывающий домен (RBD), который связывается с эффекторным доменом Ras-GTP, высвобождая CR1 и останавливая ингибирование киназы. Остатки 234–280 содержат мотив сложный эфир форбола / DAG -связывающий цинковый палец, который участвует в стыковке мембраны B-Raf после связывания Ras.
Консервированная область 2 (CR2) обеспечивает гибкий линкер, который соединяет CR1 и CR3 и действует как шарнир.
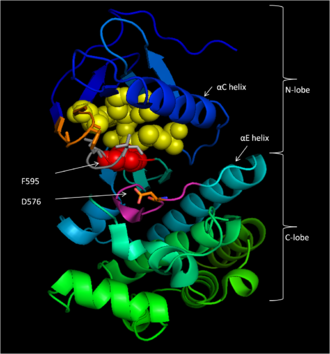 Рисунок 1: Неактивная конформация киназы B-Raf (CR3) домен. С помощью палочек показаны гидрофобные взаимодействия P-петли (оранжевый) с остатками активационной петли (серый), которые стабилизируют конформацию неактивной киназы. F595 (красный) блокирует гидрофобный карман, в котором связывается АТФ-аденин (желтый). D576 (оранжевый) показан как часть каталитического контура (пурпурный). Рисунок изменен на основе PDB id 1UWH.
Рисунок 1: Неактивная конформация киназы B-Raf (CR3) домен. С помощью палочек показаны гидрофобные взаимодействия P-петли (оранжевый) с остатками активационной петли (серый), которые стабилизируют конформацию неактивной киназы. F595 (красный) блокирует гидрофобный карман, в котором связывается АТФ-аденин (желтый). D576 (оранжевый) показан как часть каталитического контура (пурпурный). Рисунок изменен на основе PDB id 1UWH. Консервативная область 3 (CR3), остатки 457–717, составляет ферментативный киназный домен B-Raf. Эта в значительной степени консервативная структура является двулопастной, соединенной короткой шарнирной областью. Меньшая часть N (остатки 457–530) в первую очередь отвечает за связывание АТФ, в то время как большая часть C (остатки 535-717) связывает субстрат белки. Активный сайт представляет собой щель между двумя долями, а каталитический остаток Asp 576 расположен на С-доле, обращенной внутрь этой щели.
P- Петля
P-петля B-Raf (остатки 464–471) стабилизирует непереносимые фосфатные группы АТФ во время связывания фермента с АТФ. В частности, S 467, F 468 и G 469 основная цепь амиды связывают водородную связь с β-фосфатом АТФ, чтобы закрепить молекула. Функциональные мотивы B-Raf были определены путем анализа гомологии PKA, проанализированной Хэнксом и Хантером, с доменом киназы B-Raf.
Карман для связывания нуклеотидов
V 471, C 532, W 531, T 529, L 514 и A 481 образуют гидрофобный карман, внутри которого аденин АТФ закрепляется за счет притяжения Ван-дер-Ваальса при связывании АТФ.
Каталитическая петля
Остатки 574–581 составляют часть киназного домена, ответственного за поддержку переноса γ-фосфата АТФ на белковый субстрат B-Raf. В частности, D 576 действует как акцептор протона, активируя нуклеофильный гидроксильный кислород на остатках серина или треонина субстрата, позволяя протекать реакции переноса фосфата, опосредованной основной катализ.
DFG Motif
D594, F595 и G596 составляют мотив, центральный для функции B-Raf как в его неактивном, так и в активном состоянии. В неактивном состоянии F595 занимает карман для связывания нуклеотидов, препятствуя проникновению АТФ и уменьшая вероятность ферментативного катализа. В активном состоянии D594 хелатирует двухвалентный катион магния катион, который стабилизирует β- и γ-фосфатные группы АТФ, ориентируя γ-фосфат для переноса.
Петля активации
Остатки 596–600 образуют сильные гидрофобные взаимодействия с P-петлей в неактивной конформации киназы, блокируя киназу в неактивном состоянии до тех пор, пока петля активации фосфорилируется, дестабилизируя эти взаимодействия наличием отрицательного заряда. Это вызывает переход киназы в активное состояние. В частности, L597 и V600 петли активации взаимодействуют с G466, F468 и V471 P-петли, чтобы сохранить киназный домен в неактивном состоянии до тех пор, пока он не будет фосфорилирован.
B-Raf является серин / треонин-специфическая протеинкиназа. По существу, он катализирует фосфорилирование сериновых и треониновых остатков в консенсусной последовательности целевых белков с помощью АТФ, давая АДФ и фосфорилированный белок в качестве продуктов. Поскольку это высоко регулируемая передача сигнала киназа, B-Raf должен сначала связать Ras - GTP, прежде чем стать активным в качестве фермента. Как только B-Raf активирован, консервативное каталитическое ядро протеинкиназы фосфорилирует белковые субстраты, способствуя нуклеофильной атаке активированного субстрата серина или треонина гидроксил атом кислорода на γ-фосфат группе АТФ посредством бимолекулярного нуклеофильного замещения.
Домен киназы (CR3) человеческих киназ Raf ингибируется двумя механизмами: аутоингибирование собственным регулирующим Ras - GTP -связывающим доменом CR1 и отсутствием посттрансляционного фосфорилирования ключевые остатки серина и тирозина (S338 и Y341 для c-Raf) в шарнирной области CR2. Во время активации B-Raf белок аутоингибиторный домен CR1 сначала связывает эффекторный домен Ras-GTP с Ras-связывающим доменом CR1 (RBD), высвобождая домен CR3 киназы, как и другие члены человеческое семейство киназ Raf. Взаимодействие CR1-Ras позже усиливается за счет связывания цистеина -субдомена (CRD) CR1 с Ras и мембранными фосфолипидами. В отличие от A-Raf и C-Raf, которые должны быть фосфорилированы по гидроксилсодержащим остаткам CR2, прежде чем полностью высвободить CR1, чтобы стать активным, B-Raf постоянно фосфорилируется по CR2 S445. Это позволяет отрицательно заряженному фосфосерину немедленно отталкивать CR1 посредством стерических и электростатических взаимодействий, когда регуляторный домен не связан, освобождая CR3-киназный домен для взаимодействия с белками-субстратами.
После высвобождения автоингибирующего регуляторного домена CR1 домен CR3 киназы B-Raf должен измениться на его АТФ -связывающий активный конформер, прежде чем он сможет катализировать фосфорилирование белка. В неактивной конформации F595 мотива DFG блокирует гидрофобный аденин связывающий карман, в то время как остатки петли активации образуют гидрофобные взаимодействия с P-петлей, останавливая ATP от доступа к его сайту связывания. Когда активационная петля фосфорилируется, отрицательный заряд фосфата нестабилен в гидрофобной среде P-петли. В результате петля активации изменяет конформацию, вытягиваясь через C-долю домена киназы. В этом процессе он образует стабилизирующие взаимодействия β-листа с β6 цепью. Между тем, фосфорилированный остаток приближается к K507, образуя стабилизирующий солевой мостик, чтобы зафиксировать петлю активации на месте. Мотив DFG изменяет конформацию с петлей активации, заставляя F595 перемещаться из сайта связывания аденинового нуклеотида в гидрофобный карман, ограниченный спиралями αC и αE. Вместе DFG и движение активационной петли при фосфорилировании открывают сайт связывания АТФ . Поскольку все другие субстрат-связывающие и каталитические домены уже находятся на своих местах, фосфорилирование одной только петли активации активирует киназный домен B-Raf посредством цепной реакции, которая по существу удаляет крышку с подготовленного другим способом активного сайта.
 Рис. 2: Катализируемая основанием нуклеофильная атака остатка серин / треонинового субстрата на γ-фосфатную группу АТФ. Шаг 1: хелатирование вторичного иона магния с помощью N581 и депротонирование субстрата Ser / Thr с помощью D576. Шаг 2: нуклеофильная атака активированного гидроксила субстрата на γ-фосфат АТФ. Шаг 3: комплекс магния разрушается, и D576 депротонируется. Этап 4: высвобождение продуктов.
Рис. 2: Катализируемая основанием нуклеофильная атака остатка серин / треонинового субстрата на γ-фосфатную группу АТФ. Шаг 1: хелатирование вторичного иона магния с помощью N581 и депротонирование субстрата Ser / Thr с помощью D576. Шаг 2: нуклеофильная атака активированного гидроксила субстрата на γ-фосфат АТФ. Шаг 3: комплекс магния разрушается, и D576 депротонируется. Этап 4: высвобождение продуктов. Чтобы эффективно катализировать фосфорилирование белка посредством бимолекулярного замещения остатков серина и треонина на АДФ в качестве уходящей группы, B-Raf должен сначала связать АТФ а затем стабилизировать переходное состояние по мере переноса γ-фосфата АТФ.
B-Raf связывает АТФ, закрепляя нуклеотид аденина в неполярный карман (желтый, рис. 1) и ориентирующий молекулу посредством водородных связей и электростатических взаимодействий с фосфатными группами. Помимо фосфатного связывания P-петли и мотива DFG, описанного выше, K483 и E501 играют ключевую роль в стабилизации непереносимых фосфатных групп. Положительный заряд первичного амина K483 позволяет ему стабилизировать отрицательный заряд α- и β-фосфатных групп АТФ при связывании АТФ. Когда АТФ отсутствует, отрицательный заряд группы E501 карбоксильная уравновешивает этот заряд.
Когда АТФ связывается с доменом киназы B-Raf, D576 каталитической петли активирует гидроксильную группу субстрата, увеличивая ее нуклеофильность, чтобы кинетически управлять реакцией фосфорилирования, в то время как другие остатки каталитической петли стабилизируют переходное состояние (рис. 2). N581 хелатирует двухвалентный катион магния, связанный с АТФ, чтобы помочь сориентировать молекулу для оптимального замещения. K578 нейтрализует отрицательный заряд на γ-фосфатной группе АТФ, так что активированный остаток субстрата ser / thr не будет испытывать такое сильное электрон-электронное отталкивание при атаке фосфата. После переноса фосфатной группы высвобождаются АДФ и новый фосфопротеин.
Поскольку конститутивно активные мутанты B-Raf обычно вызывают рак (см. Клиническое значение) из-за чрезмерной передачи сигналов клеткам для роста, ингибиторы B-Raf были разработаны как для неактивной, так и для активной конформации киназного домена в качестве кандидатов для лечения рака.
 Рисунок 3: B-Raf-киназный домен заблокирован в неактивной конформации за счет связанного BAY43 -9006. Гидрофобные взаимодействия закрепляют BAY43-9006 в сайте связывания АТФ, в то время как водородная связь группы мочевины улавливает D594 мотива DFG. Трифторметил фенил кольцо BAY43-9006 дополнительно запрещает движение DFG-мотива и петли активации к активному кондеру через стерическую блокировку.
Рисунок 3: B-Raf-киназный домен заблокирован в неактивной конформации за счет связанного BAY43 -9006. Гидрофобные взаимодействия закрепляют BAY43-9006 в сайте связывания АТФ, в то время как водородная связь группы мочевины улавливает D594 мотива DFG. Трифторметил фенил кольцо BAY43-9006 дополнительно запрещает движение DFG-мотива и петли активации к активному кондеру через стерическую блокировку. BAY43-9006 (Сорафениб, Нексавар) представляет собой V600E мутант B-Raf и C-Raf ингибитор, одобренный FDA для лечения первичной печени и почек рак. Bay43-9006 отключает домен B-Raf киназы, блокируя фермент в его неактивной форме. Ингибитор выполняет это путем блокирования кармана связывания АТФ за счет высокой аффинности к киназному домену. Затем он связывает ключевую активационную петлю и остатки мотива DFG, чтобы остановить движение активационной петли и мотива DFG к активной конформации. Наконец, трифторметилфенильный фрагмент стерически блокирует мотив DFG и активный сайт конформации петли активации, делая невозможным сдвиг конформации киназного домена, чтобы он стал активным.
Дистальное пиридильное кольцо BAY43 -9006 закрепляется в гидрофобном нуклеотид-связывающем кармане N-доли киназы, взаимодействуя с W531, F583 и F595. Гидрофобные взаимодействия с каталитической петлей F583 и мотивом DFG F595 стабилизируют неактивную конформацию этих структур, снижая вероятность активации фермента. Дальнейшее гидрофобное взаимодействие K483, L514 и T529 с центральным фенильным кольцом увеличивает сродство киназного домена к ингибитору. Гидрофобное взаимодействие F595 с центральным кольцом также снижает энергетическую привлекательность переключателя конформации DFG. Наконец, полярные взаимодействия BAY43-9006 с киназным доменом продолжают эту тенденцию увеличения ферментативного сродства к ингибитору и стабилизации остатков DFG в неактивной конформации. Водородные связи E501 и C532 связывают мочевину и пиридильную группы ингибитора, соответственно, в то время как мочевина карбонил принимает водородную связь от основной цепи D594 амид
Трифторметилфенильный фрагмент закрепляет термодинамическую благоприятность неактивной конформации, когда киназный домен связан с BAY43-9006, стерически блокируя гидрофобный карман между спиралями αC и αE, который мотив DFG и петля активации будут заселяться при перемещении в свои места в активной конформации белка.
 Рисунок 4: Структуры вемурафениба (справа) и его предшественника, PLX 4720 (слева), два ингибитора активной конформации домена киназы B-Raf
Рисунок 4: Структуры вемурафениба (справа) и его предшественника, PLX 4720 (слева), два ингибитора активной конформации домена киназы B-Raf PLX4032 (вемурафениб ) представляет собой мутантный V600 ингибитор B-Raf, одобренный FDA для лечения поздней стадии меланомы. В отличие от BAY43-9006, который ингибирует неактивную форму киназного домена, вемурафениб ингибирует активную «DFG-in» форму киназы, прочно закрепляясь в сайте связывания АТФ. Ингибируя только активную форму киназы, вемурафениб избирательно подавляет пролиферацию клеток с нерегулируемым B-Raf, обычно тех, которые вызывают рак.
Так как вемурафениб отличается от своего предшественника, PLX4720, только фенилом. кольцо добавлено по фармакокинетическим причинам, действие PLX4720 эквивалентно действию вемурафениба. PLX4720 имеет хорошее сродство к сайту связывания АТФ частично потому, что его якорная область, 7-аза индол бициклический, отличается только от природного аденина, который занимает сайт в двух местах, где атомы азота были заменены на карбон. Это позволяет сохранять сильные межмолекулярные взаимодействия, такие как водородная связь N7 с C532 и водородная связь N1 с Q530. Отличное расположение в гидрофобном кармане, связывающем АТФ (C532, W531, T529, L514, A481), также увеличивает сродство связывания. Кетонный линкер, связывающий водородную связь с водой, и дифторфенил, помещающийся во второй гидрофобный карман (A481, V482, K483, V471, I527, T529, L514 и F583), вносят вклад в исключительно высокую аффинность связывания в целом. Селективное связывание с активным Raf осуществляется концевой пропильной группой, которая связывается с Raf-селективным карманом, созданным смещением спирали αC. Селективность в отношении активной конформации киназы дополнительно увеличивается за счет pH-чувствительной депротонированной группы сульфонамида, которая стабилизируется водородными связями с пептидом основной цепи NH D594 в активном состоянии. В неактивном состоянии сульфонамидная группа ингибитора вместо этого взаимодействует с основной цепью карбонил этого остатка, создавая отталкивание. Таким образом, вемурафениб связывается преимущественно с активным состоянием киназного домена B-Raf.
Мутации в гене BRAF могут вызывать заболевание двумя путями. Во-первых, мутации могут передаваться по наследству и вызывать врожденные дефекты. Во-вторых, мутации могут появиться в более позднем возрасте и вызвать рак, поскольку онкоген.
Унаследованные мутации в этом гене вызывают кожно-сердечно-сосудистый синдром, заболевание, характеризующееся пороками сердца, умственной отсталостью и характерным внешним видом лица..
Мутации в этом гене были обнаружены при раковых заболеваниях, включая неходжкинскую лимфому, колоректальный рак, злокачественную меланому, папиллярная карцинома щитовидной железы, немелкоклеточная карцинома легкого, аденокарцинома легкого, опухоли головного мозга, включая глиобластому и плеоморфная ксантоастроцитома, а также воспалительные заболевания, такие как болезнь Эрдхейма-Честера.
Мутация V600E гена BRAF была связана с лейкемией волосатых клеток во многих исследованиях и была предложена для использования при скрининге синдрома Линча для уменьшения количества пациентов, подвергающихся ненужному секвенированию MLH1.
Более 30 измененных ионы гена BRAF, связанные с раком человека, были идентифицированы. Частота мутаций BRAF широко варьируется при раке человека, от более 80% в меланомах и невусах до всего 0–18% в других опухолях, например, 1–3% при раке легких и 5% при колоректальном раке. В 90% случаев тимин заменяется аденином в нуклеотиде 1799. Это приводит к замене валина (V) на глутамат (E) в кодоне 600 (теперь обозначаемом как V600E ) при активации сегмент, который был обнаружен при раке человека. Эта мутация широко наблюдалась при папиллярной карциноме щитовидной железы, колоректальном раке, меланоме и немелкоклеточном раке легкого. Мутация BRAF-V600E присутствует у 57% пациентов с гистиоцитозом клеток Лангерганса. Мутация V600E является вероятной драйверной мутацией в 100% случаев лейкемии волосатых клеток. Высокая частота мутаций BRAF V600E была обнаружена в амелобластоме, доброкачественном, но локально инфильтративном одонтогенном новообразовании. Мутация V600E также может быть связана как мутация с одним драйвером (генетический «дымящийся пистолет») с определенными случаями развития папиллярной краниофарингиомы.
Другие обнаруженные мутации: R461I, I462S, G463E, G463V, G465A, G465E, G465V, G468A, G468E, N580S, E585K, D593V, F594L, G595R, L596V, T598I, V599D, V599V, V599K, V277, V599R, большинство из этих мутаций и т. Д. сгруппированы в две области: богатая глицином P-петля N-доли и сегмент активации и фланкирующие области. Эти мутации изменяют активационный сегмент из неактивного состояния в активное состояние, например, в ранее цитированной статье сообщалось, что алифатическая боковая цепь Val599 взаимодействует с фенильным кольцом Phe467 в P-петле. Ожидается, что замена гидрофобной боковой цепи Val среднего размера на более крупный и заряженный остаток, обнаруживаемый при раке человека (Glu, Asp, Lys или Arg), дестабилизирует взаимодействия, которые поддерживают мотив DFG в неактивной конформации, поэтому сегмент активации в активную позицию. В зависимости от типа мутации активность киназы в отношении MEK также может варьироваться. Большинство мутантов стимулируют повышенную активность B-Raf киназы по отношению к MEK. Однако некоторые мутанты действуют по другому механизму, потому что, хотя их активность в отношении MEK снижена, они принимают конформацию, которая активирует C-RAF дикого типа, который затем передает сигнал ERK.
Как упоминалось выше, некоторые фармацевтические фирмы разрабатывают специфические ингибиторы мутированного белка B-raf для противоопухолевого препарата использование, потому что BRAF - это хорошо изученная цель с высоким выходом. Вемурафениб (RG7204 или PLX4032) был лицензирован Управлением по контролю за продуктами и лекарствами США как Зелбораф для лечения метастатической меланомы в августе 2011 г. на основании клинических данных фазы III. Было замечено улучшение выживаемости, а также степень ответа на лечение на 53% по сравнению с 7-12% при бывшем лучшем химиотерапевтическом лечении дакарбазином. В клинических испытаниях B-Raf увеличивал вероятность выживания пациента с метастатической меланомой. Несмотря на высокую эффективность препарата, 20% опухолей все еще развивают устойчивость к лечению. У мышей 20% опухолей становятся устойчивыми через 56 дней. Хотя механизмы этой устойчивости все еще обсуждаются, некоторые гипотезы включают сверхэкспрессию B-Raf для компенсации высоких концентраций вемурафениба и предшествующую регуляцию передачи сигналов роста.
Более общие ингибиторы B-Raf включают GDC-0879, PLX-4720, Сорафениб, дабрафениб и LGX818
Было показано, что BRAF (ген) взаимодействовать с:
![]() Эта статья включает материалы общественного достояния из документа Национального института рака США : «Словарь терминов по раку».Эта статья включает текст из Национальной медицинской библиотеки США, которая находится в общественном достоянии.
Эта статья включает материалы общественного достояния из документа Национального института рака США : «Словарь терминов по раку».Эта статья включает текст из Национальной медицинской библиотеки США, которая находится в общественном достоянии.
.