
Войти
| Аквапорин | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Кристаллографическая структура аквапорина 1 (AQP1 ) PDB 1j4n Кристаллографическая структура аквапорина 1 (AQP1 ) PDB 1j4n | |||||||||
| Идентификаторы | |||||||||
| Symbol | Aquaporin | ||||||||
| Pfam | PF00230 | ||||||||
| InterPro | IPR000425 | ||||||||
| PROSITE | PDOC00193 | ||||||||
| SCOPe | 1fx8 / SUPFAM | ||||||||
| TCDB | 1.A.8 | ||||||||
| суперсемейство OPM | 7 | ||||||||
| белок OPM | 2zz9 | ||||||||
| |||||||||
Аквапорины, также называемые водные каналы, являются канальными белками из более крупного семейства основных внутренних белков, которые образуют поры в мембране биологических клеток, в основном облегчая транспорт воды между ячейками. Клеточные мембраны различных бактерий, грибов, животных и клеток растений содержат аквапорины, через которые вода может быстрее течь в и из клетки, чем путем диффузии через фосфолипидный бислой . Аквапорины имеют шесть охватывающих мембрану альфа-спиральных доменов с карбоксильными и аминными окончаниями на цитоплазматической стороне. Две гидрофобные петли содержат консервативный мотив аспарагин-пролин-аланин NPA, который формирует цилиндр, окружающий центральную пористую область, которая содержит дополнительную плотность белка. Поскольку аквапорины обычно всегда открыты и преобладают практически в каждом типе клеток, это приводит к неправильному представлению о том, что вода легко проходит через клеточную мембрану вниз по градиенту ее концентрации. Вода может проходить через клеточную мембрану посредством простой диффузии, потому что это малая молекула, и посредством осмоса, в тех случаях, когда концентрация воды вне клетки выше, чем концентрация воды в ней. внутри. Однако, поскольку вода является полярной молекулой, этот процесс простой диффузии является относительно медленным, и большая часть воды проходит через аквапорин.
Нобелевская премия по химии 2003 г. был награжден совместно Питеру Агре за открытие аквапоринов и Родерику Маккиннону за его работу по структуре и механизму калиевых каналов.
Генетические дефекты, связанные с аквапорином гены были связаны с несколькими заболеваниями человека, включая нефрогенный несахарный диабет и оптический нейромиелит.
Механизм облегченного переноса воды и вероятное существование водных пор привлекают исследователей с тех пор, как 1957. В большинстве клеток вода входит и выходит за счет осмоса через липидный компонент клеточных мембран. Из-за относительно высокой проницаемости для воды некоторых эпителиальных клеток долгое время предполагалось, что должен существовать некий дополнительный механизм переноса воды через мембраны. Соломон и его сотрудники выполнили новаторскую работу по изучению проницаемости воды через клеточную мембрану в конце 1950-х годов. В середине 1960-х альтернативная гипотеза («модель разделения-диффузии») пыталась установить, что молекулы воды распределяются между водной фазой и липидной фазой, а затем диффундируют через мембрану, пересекая ее до следующей межфазной границы, где они покидают мембрану. липид и вернулся в водную фазу. Исследования Паризи, Эдельмана, Карвуниса и др. подчеркнули не только важность наличия водных каналов, но и возможность регулирования их проницаемости. В 1990 году эксперименты Веркмана продемонстрировали функциональную экспрессию водных каналов, указывая на то, что водные каналы фактически являются белками.
Только в 1992 году первый аквапорин, «аквапорин-1» (первоначально известный как CHIP 28), о чем сообщил Питер Агре из Университета Джона Хопкинса. В 1999 году вместе с другими исследовательскими группами Агре сообщил о первых изображениях с высоким разрешением трехмерной структуры аквапорина, а именно аквапорина-1. Дальнейшие исследования с использованием моделирования суперкомпьютера определили путь воды, когда она движется через канал, и продемонстрировали, как пора может пропускать воду без прохождения мелких растворенных веществ. Новаторское исследование и последующее открытие водных каналов Агре и его коллегами принесло Агре Нобелевскую премию по химии в 2003 году. Агре сказал, что он открыл аквапорины «по интуиции». Он изучал группу крови Rh антигены и выделил молекулу Rh, но вторая молекула размером 28 килодальтон (и поэтому называлась 28K) продолжала появляться. Сначала они подумали, что это фрагмент молекулы Rh или примесь, но оказалось, что это новый тип молекулы с неизвестной функцией. Он присутствует в структурах, таких как почечные канальцы и красные кровяные тельца, и связан с белками различного происхождения, такими как мозг плодовой мушки, бактерии, хрусталик глаза и ткани растений
.Однако первое сообщение об опосредованном белком переносе воды через мембраны было сделано Георге Бенга и другими в 1986 году, до первой публикации Агре по этой теме. Это привело к спорам о том, что работа Бенги не была должным образом признана ни Агре, ни Комитетом по Нобелевской премии.
 Иллюстрация молекулы аквапорина
Иллюстрация молекулы аквапорина Аквапорины - «водопроводная система для клеток». Вода движется через клетки организованным образом, наиболее быстро в тканях, которые имеют водные каналы аквапорина. В течение многих лет ученые предполагали, что вода просачивается через клеточную мембрану, и некоторое количество воды это происходит. Однако это не объясняет, как вода может так быстро перемещаться через некоторые клетки.
Аквапорины избирательно проводят молекулы воды в клетку и из нее, предотвращая прохождение ионов и других растворенных веществ. Аквапорины, также известные как водные каналы, являются составными белками пор мембраны. Некоторые из них, известные как акваглицеропорины, также переносят другие небольшие незаряженные растворенные молекулы, включая аммиак, CO 2, глицерин и мочевину. Например, канал аквапорина 3 имеет ширину пор 8–10 Ангстремов и позволяет проходить гидрофильным молекулам в диапазоне от 150 до 200 Да. Однако поры воды полностью блокируют ионы, в том числе протоны, что необходимо для сохранения разности электрохимических потенциалов мембраны.
Молекулы воды проходят через поры канала одним файлом. Наличие водных каналов увеличивает проницаемость мембраны для воды. Они также важны для водной транспортной системы растений и устойчивости к засухе и солевым стрессам.
 Схематическая диаграмма 2D-структуры аквапорина 1 (AQP1 ), изображающая шесть трансмембранные альфа-спирали и пять областей межспиральной петли AE
Схематическая диаграмма 2D-структуры аквапорина 1 (AQP1 ), изображающая шесть трансмембранные альфа-спирали и пять областей межспиральной петли AE  Трехмерная структура аквапорина Z, выделяющая водный канал в форме «песочных часов», который прорезает центр белка
Трехмерная структура аквапорина Z, выделяющая водный канал в форме «песочных часов», который прорезает центр белка Белки аквапорина состоят из пучка из шести трансмембранные α-спирали. Они встроены в клеточную мембрану. Амино- и карбоксильные концы обращены внутрь клетки. Амино- и карбоксильная половины похожи друг на друга, по-видимому, повторяя последовательность нуклеотидов. Некоторые исследователи полагают, что это было создано удвоением гена, который раньше был наполовину меньше. Между спиралями находятся пять участков (A - E), которые входят в клеточную мембрану или из нее, две из них гидрофобные (B, E), с аспарагином - пролином - образец аланина («мотив NPA»). Они создают характерную форму песочных часов, делая водяной канал узким в середине и шире на каждом конце.
Еще одно, еще более узкое место в канале - это «фильтр селективности ar / R», кластер аминокислот, позволяющий аквапорину избирательно пропускать или блокировать проход различных молекул.
Аквапорины образуют четырехкомпонентные кластеры (тетрамеры) в клеточной мембране, при этом каждый из четырех мономеров действует как водный канал. У разных аквапоринов есть водные каналы разного размера, самые маленькие из них не пропускают ничего, кроме воды.
Рентгеновские профили показывают, что аквапорины имеют два конических входа. Эта форма песочных часов может быть результатом процесса естественного отбора в сторону оптимальной проницаемости. Было показано, что конические входы с подходящим углом раскрытия действительно могут обеспечить большое увеличение проницаемости гидродинамических каналов.
Аквапориновые каналы, по-видимому, в моделировании позволяют проходить только воде, поскольку молекулы эффективно выстраиваются в очередь в один файл. Управляемый локальным электрическим полем аквапорина, кислород в каждой молекуле воды обращен вперед, когда входит, поворачиваясь на полпути и уходя, когда кислород обращен назад. Почему происходит это вращение, пока не совсем понятно. Некоторые исследователи назвали причиной электростатическое поле, создаваемое двумя полуспиралями аквапорина HB и HE. Другие предположили, что водородные связи между аминокислотами аспарагин в двух областях NPA и кислородом в воде вызывают вращение. Пока неясно, имеет ли вращение молекул воды какое-либо биологическое значение. Ранние исследования предполагали, что «биполярная» ориентация молекул воды блокирует поток протонов через механизм Гроттуса. Более поздние исследования ставят под сомнение эту интерпретацию и подчеркивают, что электростатический барьер является причиной блокировки протонов. Согласно последней точке зрения, вращение молекул воды является лишь побочным эффектом электростатического барьера. В настоящее время (2008 г.) происхождение электростатического поля является предметом дискуссий. В то время как некоторые исследования в основном рассматривали электрическое поле, создаваемое полусухими белками HB и HE, другие подчеркивали эффекты десольватации, когда протон входит в узкую пору аквапорина.
 Схематическое изображение движения воды через узкий фильтр селективности канала аквапорина
Схематическое изображение движения воды через узкий фильтр селективности канала аквапорина Фильтр селективности ароматический / аргинин или «ar / R» представляет собой кластер аминокислот, который помогают связываться с молекулами воды и исключить другие молекулы, которые могут попытаться проникнуть в поры. Это механизм, с помощью которого аквапорин может избирательно связывать молекулы воды (следовательно, пропуская их) и предотвращать проникновение других молекул. Фильтр ar / R состоит из двух аминокислотных групп спиралей B (HB) и E (HE) и двух групп петли E (LE1, LE2) с двух сторон мотива NPA. Его обычное положение - 8 Å на внешней стороне мотива NPA; обычно это самая узкая часть канала. Его узость ослабляет водородные связи между молекулами воды, позволяя аргининам, несущим положительный заряд, взаимодействовать с молекулами воды и отфильтровывать нежелательные протоны.
Существует тринадцать известных типов аквапоринов у млекопитающих, шесть из которых расположены в почках, но существует подозрение на существование гораздо большего числа. Наиболее изученные аквапорины сравниваются в следующей таблице:
| Тип | Местоположение | Функция |
|---|---|---|
| Аквапорин 1 | реабсорбция воды | |
| аквапорин 2 | реабсорбция воды в ответ на АДГ | |
| аквапорин 3 | реабсорбция воды и проницаемость глицерина | |
| аквапорин 4 | реабсорбция воды |
У растений вода забирается из почвы через корни, где она проходит из коры головного мозга в сосудистые ткани. Есть три пути, по которым вода течет в этих тканях, известных как апопластический, симпластический и трансцеллюлярный пути. В частности, аквапорины обнаруживаются в вакуолярной мембране в дополнение к плазматической мембране растений; трансцеллюлярный путь включает транспортировку воды через плазматическую и вакуолярную мембраны. Когда корни растений подвергаются воздействию хлорида ртути, который, как известно, ингибирует аквапорины, поток воды значительно уменьшается, а поток ионов - нет, что подтверждает мнение о существовании механизма переноса воды, независимого от транспорт ионов: аквапорины.
В дополнение к поддержанию нормальной осмолярности цитозоля, аквапорины могут играть важную роль в расширенном росте, обеспечивая приток воды в расширяющиеся клетки - процесс, необходимый для поддержания развития растений.
Растение аквапорины также важны для минерального питания и детоксикации ионов, которые необходимы для гомеостаза бора, кремния, мышьяка и бикарбоната.
Аквапорины в растениях разделены на пять основных гомологичных подсемейств или групп:
Эти пять подсемейств позже были разделены на более мелкие эволюционные подгруппы на основе их ДНК последовательность. PIP объединяются в две подгруппы, PIP1 и PIP2, а TIP - в 5 подгрупп: TIP1, TIP2, TIP3, TIP4 и TIP5. Каждая подгруппа снова делится на изоформы, например. ПИП1; 1, ПИП1; 2. Поскольку номенклатура изоформ исторически основана на функциональных параметрах, а не на эволюционных параметрах, несколько новых предположений о растительных аквапоринах возникли в результате изучения эволюционных отношений между различными аквапоринами. В пределах разнообразного набора изоформ аквапоринов в растениях также существуют уникальные образцы экспрессии, специфичной для клеток и тканей.
Когда растительные аквапорины подавляются, гидравлическая проводимость и фотосинтез листа
Когда происходит закрытие растительных аквапоринов, это останавливает поток воды через поры белка. Это может произойти по ряду причин, например, когда растение содержит небольшое количество клеточной воды из-за засухи. Стробирование аквапорина осуществляется за счет взаимодействия между запирающим механизмом и аквапорином, которое вызывает трехмерное изменение белка, так что он блокирует поры и, таким образом, препятствует потоку воды через поры. У растений существует по крайней мере две формы аквапоринирования: стробирование путем дефосфорилирования определенных остатков серина в ответ на засуху и протонирование определенных остатков гистидина в ответ на затопление. Фосфорилирование аквапорина участвует в открытии и закрытии лепестков в зависимости от температуры.
некоторые бактерии и многие другие организмы также экспрессируют аквапорины.
Аквапорины также были обнаружены у грибов - Saccharomyces cerevisiae (дрожжи), Dictyostelium, Candida и Ustilago и простейшие - Trypanosoma и Plasmodium.
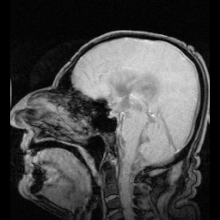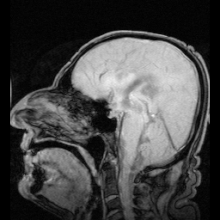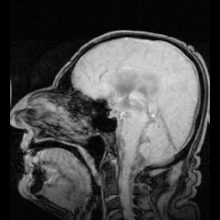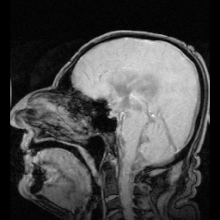 Аквапорин-1 (AQP1) обнаружен в сосудистом сплетении, способствует выработке CSF и обнаружен AQP4 на периваскулярные и эпендимные клетки.
Аквапорин-1 (AQP1) обнаружен в сосудистом сплетении, способствует выработке CSF и обнаружен AQP4 на периваскулярные и эпендимные клетки. Существуют два ярких примера заболеваний, идентифицированных как результат мутаций в аквапоринах:
Небольшое количество людей были идентифицирован с серьезным или полным дефицитом аквапорина-1. В целом они здоровы, но обнаруживают нарушение способности концентрировать растворенные вещества в моче и сохранять воду при отсутствии питьевой воды. У мышей с целевыми делециями в аквапорине-1 также наблюдается дефицит водосбережения из-за неспособности концентрировать растворенные вещества в мозговом веществе почек за счет противоточного размножения.
Помимо своей роли в генетически детерминированном несахарном почечном диабете, аквапорины также играют ключевая роль в приобретенных формах несахарного нефрогенного диабета (расстройства, вызывающие повышенное образование мочи). Аквапорин 2 регулируется вазопрессином, который при связывании с рецептором на поверхности клетки активирует сигнальный путь цАМФ. В результате аквапорин-2, содержащий везикулы, увеличивает поглощение воды и возвращается в циркуляцию. Мутация рецептора вазопрессина аквапорина 2 является причиной приобретенного несахарного диабета. У крыс приобретенный несахарный нефрогенный диабет может быть вызван нарушением регуляции аквапорина-2 из-за введения солей лития, низких концентраций калия в крови (гипокалиемия ) и высокого кальция. концентрации в крови (гиперкальциемия ).
Аутоиммунные реакции против аквапорина 4 у людей вызывают болезнь Девика. Если аквапорином можно манипулировать, это потенциально может решить медицинские проблемы, такие как задержка жидкости при сердечных заболеваниях и отек мозга после инсульта.
| На Викискладе есть материалы, связанные с аквапоринами. |