
Войти

 Два суккулентных растения рода, Euphorbia и Astrophytum, только отдаленно связаны между собой, но виды внутри каждого сошлись в сходной форме тела.
Два суккулентных растения рода, Euphorbia и Astrophytum, только отдаленно связаны между собой, но виды внутри каждого сошлись в сходной форме тела.
Конвергентная эволюция - это независимая эволюция сходных черт у видов разных периодов или эпох во времени. Конвергентная эволюция создает аналогичные структуры, которые имеют похожую форму или функцию, но не присутствуют в последнем общем предке этих групп. кладистический термин для того же явления - гомоплазия. повторяющаяся эволюция полета является классическим примером: летающие насекомые, птицы, птерозавры и летучие мыши самостоятельно развили полезную способность полета. Функционально похожие признаки, возникшие в результате конвергентной эволюции, аналогичны, тогда как гомологичные структуры или признаки имеют общее происхождение, но могут иметь разные функции. Птица, летучая мышь и птерозавр крылья являются аналогичными структурами, но их передние конечности гомологичны, разделяя наследственное состояние, несмотря на выполнение разных функций.
Противоположностью конвергенции является дивергентная эволюция, когда родственные виды развивают разные черты. Конвергентная эволюция аналогична параллельной эволюции, которая происходит, когда два независимых вида развиваются в одном направлении и, таким образом, независимо приобретают сходные характеристики; например, летающие лягушки произошли параллельно от нескольких типов древесных лягушек.
. Известно много примеров конвергентной эволюции у растений, включая повторное развитие C4фотосинтез, распространение семян мясистыми плодами, приспособленными для употребления в пищу животными, и плотоядность.
 Гомология и аналогия у млекопитающих и насекомых: на горизонтальной оси структуры гомологичны по морфологии, но различаются по функциям из-за различий в место обитания. На вертикальной оси структуры аналогичны по функциям из-за сходного образа жизни, но анатомически различаются с разной филогенией.
Гомология и аналогия у млекопитающих и насекомых: на горизонтальной оси структуры гомологичны по морфологии, но различаются по функциям из-за различий в место обитания. На вертикальной оси структуры аналогичны по функциям из-за сходного образа жизни, но анатомически различаются с разной филогенией.В морфологии аналогичные черты возникают, когда разные виды живут одинаково и / или в аналогичной среде, и поэтому сталкиваются с те же факторы окружающей среды. Занимая похожие экологические ниши (то есть особый образ жизни), аналогичные проблемы могут привести к аналогичным решениям. Британский анатом Ричард Оуэн был первым, кто идентифицировал фундаментальное различие между аналогиями и гомологиями.
В биохимии физические и химические ограничения на механизмы вызвали некоторые структуры активного центра, такие как каталитическая триада, чтобы независимо развиваться в отдельных суперсемействах ферментов.
В его книге 1989 года Wonderful Life, Стивен Джей Гулд утверждал, что если бы можно было «перемотать ленту жизни [и] те же самые условия встретились бы снова, эволюция могла бы пойти совершенно другим курсом». Саймон Конвей Моррис оспаривает этот вывод, утверждая, что конвергенция является одним из доминирующей силой в эволюции, и, учитывая, что действуют те же экологические и физические ограничения, жизнь неизбежно будет развиваться в направлении «оптимального» плана тела, и в какой-то момент эволюция обязательно наткнется на интеллект, черта, которая в настоящее время идентифицируется по крайней мере с приматы, врановые и китообразные.
В кладистике гомоплазия - это черта, общая для двух или более таксонов по любой причине, кроме того, что они имеют общее происхождение. Таксоны, которые имеют общее происхождение, являются частью одного клады ; кладистика стремится расположить их по степени родства, чтобы описать их филогению. Таким образом, гомопластические черты, вызванные конвергенцией, являются, с точки зрения кладистики, смешивающими факторами, которые могут привести к неправильному анализу.
В некоторых случаях трудно сказать, есть ли признак был утерян, а затем вновь эволюционировал конвергентно, или ген был просто выключен, а затем снова включен позже. Такое возрожденное свойство называется атавизмом. С математической точки зрения неиспользованный ген (избирательно нейтральный ) имеет постоянно снижающуюся вероятность сохранения потенциальной функциональности с течением времени. Временной масштаб этого процесса сильно различается в зависимости от филогении; у млекопитающих и птиц существует разумная вероятность остаться в геноме в потенциально функциональном состоянии в течение примерно 6 миллионов лет.
 Эволюция при аминокислоте положение. В каждом случае левая разновидность изменяется от наличия аланина (A) в определенной позиции в протеине у гипотетического предка, и теперь там есть серин (S). Вид справа может претерпевать дивергентную, параллельную или конвергентную эволюцию в этом положении аминокислоты относительно первого вида.
Эволюция при аминокислоте положение. В каждом случае левая разновидность изменяется от наличия аланина (A) в определенной позиции в протеине у гипотетического предка, и теперь там есть серин (S). Вид справа может претерпевать дивергентную, параллельную или конвергентную эволюцию в этом положении аминокислоты относительно первого вида. Когда два вида похожи по определенному признаку, эволюция определяется как параллельная, если предки также были похожи, а если и не были, то сходились. Некоторые ученые утверждали, что существует континуум между параллельной и конвергентной эволюцией, в то время как другие утверждают, что, несмотря на некоторое совпадение, между ними все еще существуют важные различия.
Когда предковые формы не указаны или неизвестны, или диапазон из рассматриваемых признаков четко не указано, различие между параллельной и конвергентной эволюцией становится более субъективным. Например, поразительный пример сходных плацентарных и сумчатых форм описан Ричардом Докинсом в Слепой часовщик как случай конвергентной эволюции, поскольку млекопитающие на каждом континенте имели долгую историю эволюции. до исчезновения динозавров, при котором накапливаются соответствующие различия.
 Эволюционная конвергенция серина и цистеиновой протеазы к одним и тем же каталитическим триадам организация кислотно-основных-нуклеофильных соединений в различных суперсемействах протеаз. Показаны триады из субтилизина, пролилолигопептидазы, протеазы TEV и папаина.
Эволюционная конвергенция серина и цистеиновой протеазы к одним и тем же каталитическим триадам организация кислотно-основных-нуклеофильных соединений в различных суперсемействах протеаз. Показаны триады из субтилизина, пролилолигопептидазы, протеазы TEV и папаина.энзимология протеаз предоставляет некоторые из наиболее ярких примеров конвергентной эволюции. Эти примеры отражают внутренние химические ограничения ферментов, что приводит к тому, что эволюция сходится к эквивалентным растворам независимо и многократно.
Сериновые и цистеиновые протеазы используют разные функциональные группы аминокислот (спирт или тиол) в качестве нуклеофила. Чтобы активировать этот нуклеофил, они ориентируют кислотный и основной остаток в каталитической триаде. Химические и физические ограничения ферментного катализа привели к тому, что идентичные структуры триад эволюционировали независимо более чем в 20 раз в разных суперсемействах ферментов.
Треониновые протеазы используют аминокислоту треонин в качестве своего катализатора нуклеофил. В отличие от цистеина и серина, треонин является вторичным спиртом (т.е. имеет метильную группу). Метильная группа треонина сильно ограничивает возможные ориентации триады и субстрата, поскольку метил вступает в конфликт либо с основной цепью фермента, либо с гистидиновым основанием. Следовательно, большинство треониновых протеаз используют N-концевой треонин, чтобы избежать таких стерических конфликтов. Несколько эволюционно независимых суперсемейств ферментов с разными складками белка используют N-концевой остаток в качестве нуклеофила. Эта общность активного сайта, но различие в укладке белков указывает на то, что активный центр эволюционировал конвергентно в этих семьях.
Conus geographus продуцирует отличную форму инсулина, который больше похож на белковые последовательности рыбьего инсулина, чем на инсулин от более близких моллюсков, что свидетельствует о конвергентной эволюции.
У насекомых существует множество примеров конвергентной эволюции с точки зрения развития устойчивости к токсинам на молекулярном уровне. Одним из хорошо охарактеризованных примеров является эволюция аминокислотных замен в четко определенных положениях в структуре α-субъединицы Na, K-АТФазы, охватывающей 15 родов и 4 порядка. Выделены синергетические отношения между заменами Q111 и N122. Конвергентная эволюция в этом случае не зависит от типа отбора или временных рамок, в которых это может произойти, а больше связана с коэволюционными отношениями, вызывающими своего рода мягкий отбор между производящими карденолид растениями и насекомыми, которые охотятся на них. их.
Конвергенция происходит на уровне ДНК и аминокислотных последовательностей, полученных трансляцией структурных генов в белки. Исследования показали сходимость аминокислотных последовательностей у летучих мышей и дельфинов, обнаруживающих эхолокацию; среди морских млекопитающих; между гигантскими и красными пандами; и между тилацином и псовыми. Конвергенция также была обнаружена в типе некодирующей ДНК, цис-регуляторных элементов, например, в скорости их эволюции; это могло указывать на положительный отбор или ослабленный очищающий отбор.
 Дельфины и ихтиозавры сошлись во многих адаптациях для быстрого плавания.
Дельфины и ихтиозавры сошлись во многих адаптациях для быстрого плавания. плавающих животных, включая рыб, таких как сельдь, морских млекопитающих, таких как дельфинов и Ихтиозавры (мезозоя ) все сходились в одной и той же обтекаемой форме. Подобная форма и плавательные приспособления присутствуют даже у моллюсков, таких как Phylliroe. Веретенообразная форма тела (трубка, сужающаяся на обоих концах), принятая многими водными животными, является приспособлением, позволяющим им перемещаться с высокой скоростью в среде с высоким сопротивлением. Сходные формы тела встречаются у безухих тюленей и ушастых тюленей : у них все еще четыре ноги, но они сильно приспособлены для плавания.
Сумчатая фауна У Австралии и плацентарных млекопитающих Старого Света есть несколько поразительно похожих форм, образованных двумя кладами, изолированными друг от друга. Тело и особенно форма черепа тилацина (тасманский тигр или тасманский волк) сходились с таковыми у Canidae, таких как рыжая лиса, Vulpes vulpes.

рыжая лиса скелет

Черепа тилацина (слева), древесный волк ( справа)

Тилацин скелет
В качестве сенсорной адаптации эхолокация развивалась отдельно у китообразных (дельфинов и китов) и летучих мышей., но от тех же генетических мутаций.
 Глаза камеры позвоночных (слева) и головоногих (справа) развивались независимо и устроены по-разному; например, волокна зрительного нерва достигают позвоночного сетчатки спереди, создавая слепое пятно.
Глаза камеры позвоночных (слева) и головоногих (справа) развивались независимо и устроены по-разному; например, волокна зрительного нерва достигают позвоночного сетчатки спереди, создавая слепое пятно.. Одним из самых известных примеров конвергентной эволюции является глаз камеры. из головоногих (таких как кальмары и осьминоги), позвоночных (включая млекопитающих) и cnidaria (таких как медузы). Их последний общий предок имел самое большее простое светочувствительное пятно, но ряд процессов привел к прогрессивному совершенствованию глаз камеры - с одним резким отличием: глаз головоногих моллюсков «запутан» в противоположном направлении, с кровеносные и нервные сосуды поступают с тыльной стороны сетчатки, а не с передней, как у позвоночных. В результате у головоногих моллюсков отсутствует слепое пятно.
 Крылья позвоночных частично гомологичны (от передних конечностей), но аналогичны органам полета у (1) птерозавров, (2) летучие мыши, (3) птицы, эволюционировали отдельно.
Крылья позвоночных частично гомологичны (от передних конечностей), но аналогичны органам полета у (1) птерозавров, (2) летучие мыши, (3) птицы, эволюционировали отдельно. Птицы и летучие мыши имеют гомологичные конечности, потому что оба они в конечном итоге произошли от наземных четвероногих, но их механизмы полета аналогичны, поэтому их крылья являются примерами функциональной конвергенции. Обе группы имели механический полет, развивались независимо. Их крылья существенно различаются по конструкции. Крыло летучей мыши представляет собой перепонку, натянутую на четыре чрезвычайно удлиненных пальца и ноги. Профиль крыла птицы состоит из перьев, прочно прикрепленных к предплечью (локтевой кости) и сильно сросшихся костей запястья и кисти (carpometacarpus ), только с крошечными остатки двух оставшихся пальцев, каждый из которых закрепил одно перо. Итак, хотя крылья летучих мышей и птиц функционально сходятся, они не сходятся анатомически. Птицы и летучие мыши также имеют высокую концентрацию цереброзидов в коже крыльев. Это улучшает эластичность кожи, что полезно для летающих животных; у других млекопитающих концентрация гораздо ниже. Вымершие птерозавры независимо развили крылья от своих передних и задних конечностей, в то время как насекомые имеют крылья, которые развились отдельно от разных органов.
Белки-летяги и сахарные планеры очень похожи по своему строению тела, с скользящими крыльями, протянутыми между конечностями, но белки-летяги - плацентарные млекопитающие, а сахарные планеры - сумчатые, широко разделенные в пределах рода млекопитающих.
Колибри-ястреб- бабочки и колибри эволюционировали схожими схемами полета и питания.
Ротовой аппарат насекомых демонстрирует множество примеров конвергентной эволюции. Ротовые органы различных групп насекомых состоят из набора гомологичных органов, специально предназначенных для питания этой группы насекомых. Конвергентная эволюция многих групп насекомых привела от первоначальных кусочно-жевательных частей рта к другим, более специализированным, производным типам функций. К ним относятся, например, хоботок насекомых, посещающих цветы, таких как пчелы и цветочные жуки, или кусающие-сосущие ротовые части кровососущих насекомых, таких как как блохи и комары.
Противоположные большие пальцы, позволяющие хватать предметы, чаще всего ассоциируются с приматами, такими как люди, обезьяны, обезьяны, и лемуры. Противоположные большие пальцы также эволюционировали у гигантских панд, но они совершенно разные по строению, имея шесть пальцев, включая большой палец, который развивается из кости запястья полностью отдельно от других пальцев.
 |  |  |
 Несмотря на подобное осветление цвета кожи после перемещения из Африки, разные гены участвовали в европейских (слева) и восточно-азиатских (справа) линиях. Несмотря на подобное осветление цвета кожи после перемещения из Африки, разные гены участвовали в европейских (слева) и восточно-азиатских (справа) линиях. | ||
Конвергентная эволюция у людей включает в себя цвет голубых глаз и светлый цвет кожи. Когда люди мигрировали из Африки, они переместились в более северные широты с менее интенсивным солнечным светом. Им было выгодно уменьшить пигментацию кожи. Очевидно, что до разделения европейских и восточноазиатских линий произошло некоторое осветление кожи, поскольку есть некоторые генетические различия в осветлении кожи, которые являются общими для обеих групп. Однако после того, как линии разошлись и стали генетически изолированными, кожа обеих групп стала более светлой, и это дополнительное осветление было связано с различными генетическими изменениями.
| Люди | Лемуры | ||
|---|---|---|---|
 |  |  |  |
| Несмотря на схожесть внешнего вида, генетическая основа голубых глаз у людей различна, и лемуры. | |||
Лемуры и люди оба являются приматами. У предков приматов были карие глаза, как и у большинства современных приматов. Генетическая основа голубых глаз у человека подробно изучена, и о ней известно много. Это не тот случай, когда отвечает один локус гена, скажем, от коричневого до синего цвета глаз. Однако один локус отвечает примерно за 80% вариации. У лемуров различия между голубыми и карими глазами полностью не известны, но один и тот же локус гена не задействован.
 В мирмекохории семена, такие как семена Chelidonium majus имеют твердое покрытие и прикрепленное масляное тело, элайосому, для распространения муравьями.
В мирмекохории семена, такие как семена Chelidonium majus имеют твердое покрытие и прикрепленное масляное тело, элайосому, для распространения муравьями. Хотя конвергентная эволюция часто иллюстрируется примерами животных, она часто происходила в завод эволюция. Например, C4фотосинтез, один из трех основных биохимических процессов связывания углерода, возникал независимо до 40 раз. Около 7600 видов растений покрытосеменных используют углеродную фиксацию C 4, при этом многие однодольные, включая 46% злаков, таких как кукуруза и сахарный тростник и двудольные, включая несколько видов Chenopodiaceae и Amaranthaceae.
Хороший пример конвергенции у растений эволюция съедобных фруктов, таких как яблоки. Эти семечки включают в себя (пять) плодолистиков и их вспомогательные ткани, образующие сердцевину яблока, окруженные структурами снаружи ботанического плода, цветоложем или гипантием.. Другие съедобные плоды включают другие ткани растений; например, мясистая часть помидора представляет собой стенки околоплодника. Это подразумевает конвергентную эволюцию под давлением отбора, в данном случае конкуренция за распространение семян животными через потребление мясистых плодов.
Распространение семян муравьями (мирмекохория ) эволюционировало независимо более чем в 100 раз, и присутствует в более чем 11 000 видов растений. Это один из самых ярких примеров конвергентной эволюции в биологии.
 Молекулярная конвергенция у хищных растений
Молекулярная конвергенция у хищных растений Плотоядность эволюционировала несколько раз независимо у растений в сильно разделенных группах.. У трех изученных видов: Cephalotus follicularis, Nepenthes alata и Sarracenia purpurea, наблюдается конвергенция на молекулярном уровне. Плотоядные растения выделяют ферменты в производимую ими пищеварительную жидкость. Изучая ферменты фосфатаза, гликозилгидролаза, глюканаза, РНКаза и хитиназа как а также протеин, связанный с патогенезом, и протеин, родственный тауматину, авторы обнаружили множество конвергентных замен аминокислоты. Эти изменения происходили не на каталитических участках ферментов, а на открытых поверхностях белков, где они могли взаимодействовать с другими компонентами клетки или пищеварительной жидкости. Авторы также обнаружили, что гомологичные гены в нехищном растении Arabidopsis thaliana имеют тенденцию к увеличению экспрессии, когда растение подвергается стрессу, что позволяет авторам предположить, что чувствительные к стрессу белки имеют часто использовались в повторяющейся эволюции плотоядности.
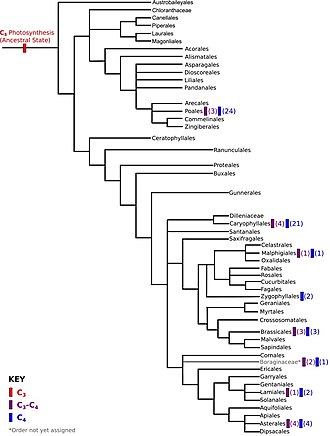 Покрытосеменные филогения отрядов, основанная на классификации по Филогенетической группе покрытосеменных. На рисунке в скобках показано количество предполагаемых независимых источников C 3-C4фотосинтеза и C4фотосинтеза.
Покрытосеменные филогения отрядов, основанная на классификации по Филогенетической группе покрытосеменных. На рисунке в скобках показано количество предполагаемых независимых источников C 3-C4фотосинтеза и C4фотосинтеза. Филогенетическая реконструкция и реконструкция предкового состояния исходят из предположения, что эволюция произошла без конвергенции. Однако конвергентные паттерны могут появляться на более высоких уровнях филогенетической реконструкции, и иногда исследователи явно ищут их. Методы, применяемые для вывода конвергентной эволюции, зависят от того, ожидается ли конвергенция на основе шаблонов или процессов. Конвергенция на основе паттернов - это более широкий термин, когда две или более линий независимо развивают паттерны схожих признаков. Конвергенция, основанная на процессах, - это когда конвергенция обусловлена аналогичными силами естественного отбора.
Ранние методы измерения конвергенции включают соотношения фенотипического и филогенетического расстояния путем моделирования эволюции с помощью модели броуновского движения эволюции черт в филогенезе. Более современные методы также позволяют количественно оценить силу сходимости. Следует иметь в виду один недостаток: эти методы могут спутать длительный застой с конвергенцией из-за фенотипического сходства. Застой возникает, когда между таксонами мало эволюционных изменений.
Измерения на основе расстояния оценивают степень сходства между линиями во времени. Частотные измерения позволяют оценить количество родословных, которые развились в определенном пространстве признаков.
Методы для вывода основанных на процессах моделей конвергенции, подходящих для выбора филогении и непрерывности данные о признаках, чтобы определить, действовали ли одни и те же селективные силы на клоны. При этом используется процесс Орнштейна-Уленбека (OU) для тестирования различных сценариев выбора. Другие методы основаны на априорной спецификации того, где произошли сдвиги в выборе.