
Войти
 | |
| Имена | |
|---|---|
| Название ИЮПАК 4-амино-5-метил-3Н-пиримидин-2-он | |
| Идентификаторы | |
| Номер CAS | |
| 3D-модель (JSmol ) | |
| 3DMet | |
| Ссылка Beilstein | 120387 |
| ChEBI | |
| ChemSpider | |
| ECHA InfoCard | 100.008.236 |
| Номер EC |
|
| KEGG | |
| MeSH | 5-Метилцитозин |
| PubChem CID | |
| UNII | |
| Панель управления CompTox (EPA ) | |
InChI
| |
УЛЫБКА
| |
| Свойства | |
| Химическая формула | C5H7N3O |
| Молярная масса | 125,131 г · моль |
| Опасности | |
| Пиктограммы GHS |  |
| Сигнальное слово GHS | Предупреждение |
| Указания на опасность GHS | H317, H319 |
| Меры предосторожности GHS | P261, P264, P272, P280, P302 + 352, P305 + 351 + 338, P321, P333 + 313, P337 + 313, P363, P501 |
| Если не указано иное отмечено, данные приведены для материалов в их стандартном состоянии (при 25 ° C [77 ° F], 100 кПа). | |
| Ссылки в ink | |
5-Метилцитозин представляет собой метилированную форму ДНК основания цитозина (C), которая регулирует транскрипцию гена и выполняет несколько других биологических ролей. Когда цитозин метилирован, ДНК сохраняет ту же последовательность, но экспрессия метилированных генов может быть изменена (изучение этого является частью области эпигенетики ). 5-Метилцитозин включен в нуклеозид 5-метилцитидин.
. В 5-метилцитозине метильная группа присоединена к 5-му атому в 6-атомном кольце, отсчет против часовой стрелки от NH азота в положении «шесть часов». Эта метильная группа отличает 5-метилцитозин от цитозина.
При попытке выделить бактериальный токсин, ответственный за туберкулез, WG Ruppel выделил новую нуклеиновую кислоту, названную туберкулиновая кислота, в 1898 году из Tubercle bacillus. Было обнаружено, что нуклеиновая кислота является необычной, поскольку она содержит, помимо тимина, гуанина и цитозина, метилированный нуклеотид. В 1925 году Джонсон и Когхилл успешно обнаружили небольшое количество метилированного производного цитозина как продукт гидролиза туберкулиновой кислоты серной кислотой. Этот отчет подвергся резкой критике, поскольку их идентификация была основана исключительно на оптических свойствах кристаллического пикрата, и другие ученые не смогли воспроизвести тот же результат. Но его существование было окончательно доказано в 1948 г., когда Хотчкисс отделил нуклеиновые кислоты ДНК от тимуса тимуса с помощью бумажной хроматографии., с помощью которого он обнаружил уникальный метилированный цитозин, совершенно отличный от обычного цитозина и урацила. Спустя семь десятилетий выяснилось, что это также общая черта в различных молекулах РНК, хотя точная роль неизвестна.
Функция этого химический состав значительно различается у разных видов:
В то время как спонтанное дезаминирование цитозина образует урацил, который распознается и удаляется ферментами репарации ДНК, дезаминирование 5-метилцитозина образует тимин. Это преобразование основания ДНК из цитозина (C) в тимин (T) может привести к переходной мутации . Кроме того, активное ферментативное дезаминирование цитозина или 5-метилцитозина семейством цитозиндезаминаз APOBEC может иметь положительное влияние на различные клеточные процессы, а также на эволюцию организма. С другой стороны, влияние дезаминирования на 5-гидроксиметилцитозин остается менее изученным.
Группа NH 2 может быть удалена (дезаминирована) из 5-метилцитозина с образованием тимина с использованием таких реагентов, как азотистая кислота ; цитозин дезаминируется до урацила (U) в аналогичных условиях.
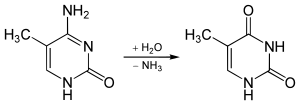 Дезаминирование 5-метилцитозина до тимина
Дезаминирование 5-метилцитозина до тимина 5-метилцитозин устойчив к дезаминированию при обработке бисульфитом, которая дезаминирует остатки цитозина. Это свойство часто используется для анализа паттернов метилирования цитозина ДНК с помощью бисульфитного секвенирования.
Метки 5mC помещаются на геномную ДНК с помощью ДНК-метилтрансфераз (ДНМТ). У человека есть 5 DNMT: DNMT1, DNMT2, DNMT3A, DNMT3B и DNMT3L, а в водорослях и грибах присутствуют еще 3 (DNMT4, DNMT5 и DNMT6). DNMT1 содержит последовательность нацеливания на фокус репликации (RFTS) и домен CXXC, которые катализируют добавление меток 5mC. RFTS направляет DNMT1 в локусы репликации ДНК, чтобы помочь в поддержании 5mC на дочерних цепях во время репликации ДНК, тогда как CXXC содержит домен цинкового пальца для добавления de novo метилирования к ДНК. Было обнаружено, что DNMT1 является преобладающей ДНК-метилтрансферазой во всех тканях человека. В первую очередь, DNMT3A и DNMT3B ответственны за метилирование de novo, а DNMT1 поддерживает метку 5mC после репликации. DNMT могут взаимодействовать друг с другом для увеличения способности метилирования. Например, 2 DNMT3L могут образовывать комплекс с 2 DNMT3A для улучшения взаимодействия с ДНК, облегчая метилирование. Изменения в экспрессии DNMT приводят к аберрантному метилированию. Сверхэкспрессия вызывает усиление метилирования, тогда как нарушение фермента снижает уровень метилирования.
 Добавление метильной группы к цитозину
Добавление метильной группы к цитозину Механизм добавления следующий: сначала остаток цистеина на мотиве PCQ DNMT создает нуклеофильную атаку на углерод 6 на цитозиновом нуклеотиде, который должен быть метилирован. S-аденозилметионин затем отдает метильную группу углероду 5. Основание в ферменте DNMT депротонирует остаточный водород на углероде 5, восстанавливая двойную связь между углеродом 5 и 6 в кольце с образованием основания 5-метилцитозина. пара.
После того, как цитозин метилирован до 5mC, он может быть возвращен обратно в исходное состояние с помощью нескольких механизмов. Пассивное деметилирование ДНК путем разбавления удаляет метку постепенно за счет репликации из-за отсутствия поддержки со стороны DNMT. При активном деметилировании ДНК серия окислений превращает ее в 5-гидроксиметилцитозин (5hmC), 5-формилцитозин (5fC) и 5-карбоксилцитозин (5caC), и последние два в конечном итоге вырезаются тиминовой ДНК-гликозилазой (TDG) с последующей эксцизионной репарацией оснований (BER) для восстановления цитозина. Нокаут TDG приводил к двукратному увеличению 5fC без каких-либо статистически значимых изменений до уровней 5hmC, что указывает на необходимость повторного окисления 5mC, по крайней мере, дважды до его полного деметилирования. Окисление происходит посредством диоксигеназ семейства TET (транслокация Ten-eleven) (ферменты TET ), которые могут превращать 5mC, 5hmC и 5fC в их окисленные формы. Однако фермент имеет наибольшее предпочтение для 5mC, и начальная скорость реакции для превращений 5hmC и 5fC с TET2 в 4,9-7,6 раз ниже. TET требует Fe (II) в качестве кофактора, а также кислорода и α-кетоглутарата (α-KG) в качестве субстратов, и последний субстрат генерируется из изоцитрата с помощью фермента изоцитратдегидрогеназа (IDH). Однако рак может продуцировать 2-гидроксиглутарат (2HG), который конкурирует с α-KG, снижая активность TET и, в свою очередь, уменьшая превращение 5mC в 5hmC.
При раке ДНК может стать как чрезмерно метилированной, называемой гиперметилированием, так и недостаточно метилированной, называемой гипометилированием. CpG-островки, перекрывающие промоторы генов, метилируются de novo, что приводит к аберрантной инактивации генов, обычно связанных с ингибированием роста опухолей (пример гиперметилирования). Сравнивая опухоль и нормальную ткань, у первой были повышенные уровни метилтрансфераз DNMT1, DNMT3A и в основном DNMT3B, все из которых связаны с аномальными уровнями 5mC при раке. Повторяющиеся последовательности в геноме, включая сателлитную ДНК, Alu и длинные вкрапленные элементы (LINE), часто наблюдаются гипометилированными при раке, что приводит к экспрессии этих обычно заглушенных генов, и уровни часто являются значимыми маркерами прогрессирования опухоли. Было высказано предположение, что существует связь между гиперметилированием и гипометилированием; Избыточная активность ДНК-метилтрансфераз, которые вызывают аномальное de novo 5mC метилирование, может быть компенсирована удалением метилирования, типом эпигенетической репарации. Однако удаление метилирования неэффективно, что приводит к перерегулированию гипометилирования всего генома. Возможно и обратное; сверхэкспрессия гипометилирования может подавляться гиперметилированием всего генома. Возможности отличительного признака рака, вероятно, приобретаются в результате эпигенетических изменений, которые изменяют 5mC как в раковых клетках, так и в окружающей опухоль-ассоциированной строме в микросреде опухоли.
«Эпигенетический возраст» «относится к связи между хронологическим возрастом и уровнями метилирования ДНК в геноме. Сочетание уровней метилирования ДНК в определенных наборах CpG, называемых «часовыми CpG», с алгоритмами, регрессирующими типичные уровни коллективного метилирования всего генома в заданном хронологическом возрасте, позволяет предсказывать эпигенетический возраст. В молодости (0–20 лет) изменения метилирования ДНК происходят более быстрыми темпами по мере развития и роста, а в более старшем возрасте изменения начинают замедляться. Существует множество эпигенетических оценок возраста. Часы Хорвата измеряют набор из 353 CpG из нескольких тканей, половина из которых положительно коррелирует с возрастом, а другая половина - отрицательно, чтобы оценить эпигенетический возраст. Часы Hannum используют образцы крови взрослых для расчета возраста на основе ортогональной основы 71 CpG. Часы Левина, известные как DNAm PhenoAge, зависят от 513 CpG и превосходят другие оценки возраста в прогнозировании смертности и продолжительности жизни, но демонстрируют систематическую ошибку с тканями, не связанными с кровью. Имеются сообщения об оценках возраста с состоянием метилирования только одного CpG в гене ELOVL2. Оценка возраста позволяет прогнозировать продолжительность жизни на основе ожиданий возрастных состояний, которым могут быть подвержены люди, на основе их маркеров метилирования 5mC.