
Войти
5-HT 3 рецептор принадлежит суперсемейство Cys-петли из лиганд-управляемых ионных каналов (LGIC) и, следовательно, отличается структурно и функционально от всех других 5-HT рецепторов (5-гидрокситриптамин, или серотонин ) рецепторы, которые представляют собой рецепторы, связанные с G-белком. Этот ионный канал является катионным -селективным и опосредует деполяризацию и возбуждение нейронов в центральной и периферической нервной системе.
Как и в случае других лиганд-зависимых ионов. каналов, рецептор 5-HT 3 состоит из пяти субъединиц, расположенных вокруг центральной ионопроводящей поры, проницаемой для натрия (Na), калия (K) и ионов кальция (Ca). Связывание нейротрансмиттера 5-гидрокситриптамина (серотонин ) с рецептором 5-HT 3 открывает канал, что, в свою очередь, приводит к возбуждающей реакции в нейроны. Быстро активирующий, снижающий чувствительность, входящий ток преимущественно переносится ионами натрия и калия. Рецепторы 5-HT 3 обладают незначительной проницаемостью для анионов. Они наиболее тесно связаны по гомологии с никотиновым ацетилхолиновым рецептором.
5-HT 3 Рецептор заметно отличается по структуре и механизму от других подтипов 5-HT рецепторов, которые все связаны с G-белком. Функциональный канал может состоять из пяти идентичных 5-HT 3A субъединиц (гомопентамерных) или смеси 5-HT 3A и одной из четырех других 5-HT 3B, 5-HT 3C, 5-HT 3D или 5-HT 3E субъединицы (гетеропентамерные). По-видимому, только субъединицы 5-HT 3A образуют функциональные гомопентамерные каналы. Все другие подтипы субъединиц должны гетеропентамеризоваться с субъединицами 5-HT 3A для образования функциональных каналов. Кроме того, в настоящее время не обнаружено каких-либо фармакологических различий между гетеромерным 5-HT 3AC, 5-HT 3AD, 5-HT 3AE и гомомерным 5-HT 3A рецептор. N-концевое гликозилирование субъединиц рецептора имеет решающее значение для сборки субъединиц и транспортировки через плазматическую мембрану.
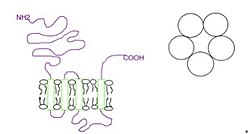 Рисунок 2. Субъединицы собраны в виде пентамера (справа), и каждая субъединица имеет четыре трансмембранных домена (слева).
Рисунок 2. Субъединицы собраны в виде пентамера (справа), и каждая субъединица имеет четыре трансмембранных домена (слева). Субъединицы окружают центральный ионный канал псевдосимметричным образом (рис.1). Каждая субъединица включает внеклеточный N-концевой домен, который включает сайт связывания ортостерического лиганда; трансмембранный домен, состоящий из четырех взаимосвязанных альфа-спиралей (M1-M4), с внеклеточной петлей M2-M3, участвующей в механизме стробирования; большой цитоплазматический домен между M3 и M4, участвующий в переносе и регуляции рецепторов; и короткий внеклеточный С-конец (фиг.1). В то время как внеклеточный домен является местом действия агонистов и конкурентных антагонистов, трансмембранный домен содержит центральную ионную пору, рецепторные ворота и основной фильтр селективности, который позволяет ионы, пересекающие клеточную мембрану.
Гены, кодирующие человеческие 5-HT 3 рецепторы, расположены на хромосомах 11 (HTR3A, HTR3B) и 3 (HTR3C, HTR3D, HTR3E), таким образом, похоже, что они возникли из дупликаций гена. Гены HTR3A и HTR3B кодируют субъединицы 5-HT 3A и 5-HT 3B и HTR3C, HTR3D и HTR3E кодируют субъединицы 5-HT 3C, 5-HT 3D и 5-HT 3E. HTR3C и HTR3E, по-видимому, не образуют функциональных гомомерных каналов, но при совместной экспрессии с HTR3A они образуют гетеромерный комплекс со сниженной или повышенной 5-HT эффективностью. Патофизиологическая роль этих дополнительных субъединиц еще предстоит идентифицировать.
Ген рецептора 5-HT 3A человека аналогичен по структуре гену мыши, который имеет 9 экзонов и занимает площадь около 13 т.п.н. Четыре из его интронов находятся точно в том же положении, что и интроны в гене гомологичного α7-ацетилхолинового рецептора, что ясно показывает их эволюционное родство.
 Рисунок 3. Структура мыши Ген рецептора 5HT3, показывающий его 9 экзонов (E1-E9), соответствующих экзонам, показанным на кДНК ниже. 5'-концы экзонов 2, 6 и 9 имеют альтернативные сайты сплайсинга. Рисунок в масштабе. Изменено по Uetz et al. 1994.
Рисунок 3. Структура мыши Ген рецептора 5HT3, показывающий его 9 экзонов (E1-E9), соответствующих экзонам, показанным на кДНК ниже. 5'-концы экзонов 2, 6 и 9 имеют альтернативные сайты сплайсинга. Рисунок в масштабе. Изменено по Uetz et al. 1994. Выражение . Гены 5-HT 3C, 5-HT 3D и 5-HT 3E имеют тенденцию демонстрировать периферически ограниченный паттерн экспрессии с высокими уровнями в кишка. Например, в двенадцатиперстной кишке и желудке человека 5-HT 3C и 5-HT 3EмРНК может быть больше, чем для 5-HT 3A и 5-HT 3B.
Полиморфизм . У пациентов, получавших химиотерапевтические препараты, определенный полиморфизм гена HTR3B может предсказать успешное противорвотное лечение. Это может указывать на то, что субъединица 5-HTR3B рецептора может использоваться в качестве биомаркера эффективности противорвотного лекарственного средства.
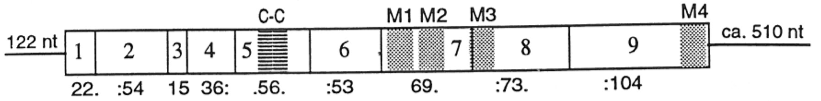 Фигура 4. Последовательность кДНК 5HT3 рецептора мыши . КДНК кодирует 122 нуклеотидную 5 'UTR и ~ 510 нуклеотидных 3' UTR. Прямоугольники указывают экзоны, а числа под экзонами указывают их длину. Например, первый экзон кодирует 22 аминокислоты плюс один нуклеотид, принадлежащий разделенному кодону, а еще 2 нуклеотида кодируются следующим экзоном. M1-4 обозначает трансмембранные спирали, а C-C обозначает петлю цистеина. Изменено по Uetz et al. 1994
Фигура 4. Последовательность кДНК 5HT3 рецептора мыши . КДНК кодирует 122 нуклеотидную 5 'UTR и ~ 510 нуклеотидных 3' UTR. Прямоугольники указывают экзоны, а числа под экзонами указывают их длину. Например, первый экзон кодирует 22 аминокислоты плюс один нуклеотид, принадлежащий разделенному кодону, а еще 2 нуклеотида кодируются следующим экзоном. M1-4 обозначает трансмембранные спирали, а C-C обозначает петлю цистеина. Изменено по Uetz et al. 1994 Рецептор 5-HT 3 экспрессируется в центральной и периферической нервной системе и опосредует различные физиологические функции. На клеточном уровне было показано, что постсинаптические 5-HT 3 рецепторы опосредуют быструю возбуждающую синаптическую передачу в неокортикальных интернейронах крыс, миндалине и гиппокампе, а также у хорьков зрительно. кора. Рецепторы 5-HT 3 также присутствуют на пресинаптических нервных окончаниях. Имеются некоторые данные о роли в модуляции высвобождения нейромедиаторов, но данные неубедительны.
Когда рецептор активируется для открытия ионного канала агонистами, наблюдаются следующие эффекты:
Агонисты рецептора включают:
Антагонисты рецептора (отсортированные по их соответствующему терапевтическому применению) включают:
Эти агенты не являются агонистами рецептора, но увеличивают сродство или эффективность рецепторов в отношении агониста:
Идентификация 5- Рецептор HT 3 не применялся до 1986 г., не имея селективных фармакологических все инструменты. Однако с открытием того, что рецептор 5-HT 3 играет важную роль в химиотерапии - и радиотерапии -индуцированной рвоте, и Сопутствующая разработка селективных антагонистов рецептора 5-HT 3 для подавления этих побочных эффектов вызвала большой интерес со стороны фармацевтической промышленности и, следовательно, идентификация 5-HT 3 быстро последовали рецепторы в клеточных линиях и нативных тканях.